




















































Tyrannosaurus

This illustration is available as a printable colouring sheet. Just click here and right click on the image that opens in a new window and save to your computer.
Name:
Tyrannosaurus
(Tyrant lizard).
Phonetic: Ty-ran-no-sore-us
Named By: Henry Fairfield Osborn - 1905.
Synonyms: Dinotyrannus megagracilis,
Dynamosaurus imperiosus, Manospondylus gigas, Nanotyrannus?,
Stygivenator molnari.
Classification: Chordata, Reptilia, Dinosauria,
Saurischia, Theropoda, Tyrannosauria, Tyrannosauridae.
Species: T. rex (type)
Type: Carnivore.
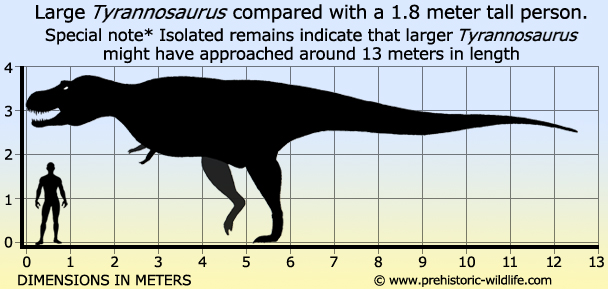
Size: Most complete specimen 12.3 meters long,
isolated remains suggest a potential upper size of around 13 meters
long.
Known locations: Western North America.
Time period: Late Campanian/Early Maastrichtian of
the Cretaceous.
Fossil representation: Over 30 individuals are
known, some forming almost complete skeletons.
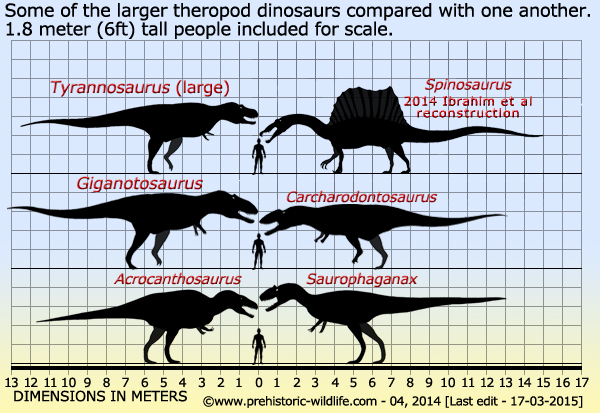
Tyrannosaurus rex has been known to science since the end of the nineteenth century, and to this day remains arguably the most popular dinosaur the world over. This is evidenced by its almost mandatory inclusion in books, TV shows, games and websites that are about dinosaurs in general. Tyrannosaurus also probably has more toys and merchandising relating to it than any other dinosaur.
Size and Growth
Although
Tyrannosaurus has lost the epithet of largest
carnivorous dinosaur to
others like Giganotosaurus
and Spinosaurus,
it still remains the
largest from North America, and still sits among the biggest known
carnivorous theropods. The largest recovered specimen of
Tyrannosaurus is an 85% complete individual
nicknamed 'Sue'
after its discoverer, and measures a total length of 12.3
meters.
However
the largest Tyrannosaurus skull measures 1.5
meters long, bigger
than 'Sue's' skull that measures 1.41 meters long. This Skull
is 6 per cent bigger than 'Sue', and by scaling up
'Sue's'' body length by the same percentage, gives a potential
total body size of up to 13 meters long. It should be
remembered however that the maximum size of Tyrannosaurus,
and indeed
other extinct creatures, needs to be attributed to fossil material of
a size that fits the estimate, otherwise it can only remain as
speculation.
Not
only is 'Sue' 12.3 meters long, study of the fossils has
revealed that 'Sue' was 28 years old at the time of death.
This is considered to be old for a Tyrannosaurus,
with maximum age
estimates for the genus usually stopping at 30 years old. It
could still be that Tyrannosaurus potentially grew
bigger than the
confirmed 12.3 meters long by the way it grew up.
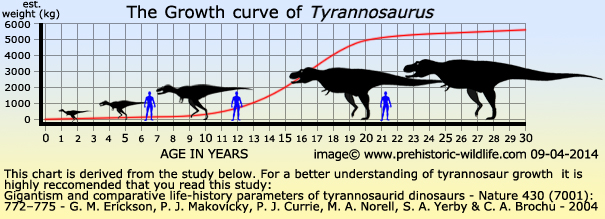
There
are a large number of fossils not just for Tyrannosaurus,
but for the
tyrannosaurid group as a whole, that represent individuals of
different ages and their corresponding sizes. When these sizes and
ages are put together, an S shaped growth curve is clearly
displayed. This indicates that for roughly the first third of their
lives, the tyrannosaurids grew quite slowly, reaching about a tenth
of their full potential size. When they became teenagers however,
they underwent a massive growth spurt, reaching nearly their full
adult size in only about ten years. After this spurt, the rate of
growth levels off as dramatically as it began when the individual
reaches approximately twenty years of age.
While
this is a common trait for the tyrannosaurid group, the rate of
growth for Tyrannosaurus is even greater than the
smaller
tyrannosaurids like Gorgosaurus.
This is even more spectacular when
you consider that Tyrannosaurus is not thought to
have lived any longer
than its smaller brethren, but still grew meters longer, and
several tons heavier.
 |
|
|
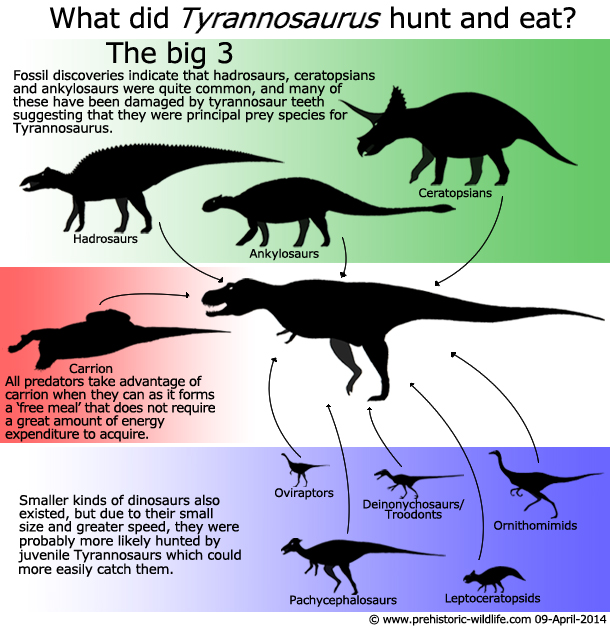
Predator or Scavenger
One
of the most controversial theories about Tyrannosaurus
is if it was
actually a predator at all, but instead lived the life of a
scavenger. Some have said that its size, even when combined with
lower weight estimates meant that it could not run, as in both feet
off the ground at the same time. Biomechanical studies have also
shown that it was no way near as fast as it is sometimes depicted in
movies. It is popularly said that for a Tyrannosaurus
to run at 25
miles per hour, it would need up to of around 80% of its
muscle mass in its legs, an unheard of scenario. What are
considered more realistic estimates place top speed at around the 11
miles per hour mark.
A
slower than expected speed does not mean that Tyrannosaurus
was a
scavenger however. You have to remember that predators need to be
only fast enough to catch their prey. Many of the primary prey
species of Tyrannosaurus were hadrosaurs
like Edmontosaurus,
ceratopsians
like Triceratops,
and maybe even
armoured dinosaurs
like Ankylosaurus.
These dinosaurs were built for browsing on
vegetation, not speed. Even though hadrosaurs are considered to
have been optionally bipedal when the situation arose, and some think
that ceratopsian dinosaurs could run fast in a similar fashion to a
rhinoceros, if they still could not out run a Tyrannosaurus
whether
through injury, old age or just inability, they would almost
certainly still be on the menu.
When
determining speed, you also have to look at the stride of the
animal, or rather, how much distance can it cover when taking a
comfortable stride forwards. If Tyrannosaurus
could out stride its
prey, it would not have to move its leg muscles as often as its prey
did, because the mechanical advantage of its longer legs would
compensate.
It's
like if you have a compass and open it up to 1 centimetres width
between the points. You place the first point on a piece of paper and
rotate it 180 degrees so the second point is in front of the
first. You then take another 'step' by rotating the first point
180 degrees around the second. 'Stepping' this way should mean
that you need to take 10 steps to clear a straight line of 10
centimetres length. Now if you repeat this, but this time use a
2 centimetre space between the compass points, you can cover the
same distance as before with just 5 'steps'. If you get two
compasses and set them to different widths, you can see the wider
compass gaining and overtaking the smaller when the number of steps is
equal.
Of
course we all know that animal legs don't work in the same way as a
rotating compass, but the principal of a larger fixed gait gaining
ground on a smaller gait still applies. As stated above, a prey
animal with a smaller stride than Tyrannosaurus
would have to move at a
faster rate in order to maintain distance.
In
determining top running speed, researchers also look at correlations
in different bone sizes. Animals that are more fleet of foot tend to
have lower leg bones (tibia and fibula) that are longer than the
upper (femur). In Tyrannosaurus these two
areas are roughly the
same size as one another, and because of this some have again
suggested that Tyrannosaurus was more likely a
scavenger as opposed to
a running predator. Again however things are not as clear cut as they
first seem.
In
Tyrannosaurus the metatarsals (foot bones) are
not positioned flat
as they are in human feet. They instead form an extension to the
lower leg bones, increasing the total length of the lower leg area.
This would be like a human being running while balancing on just their
toes instead of their whole foot, and would have worked to increase
the striding distance of Tyrannosaurus. This
arrangement of foot
bones is referred to as digitigrade, and is commonly seen in animals
that run after others.
Another
thing that supporters of the idea that Tyrannosaurus
was a scavenger
point to are the proportionately tiny arms. These have long been
conceived as being useless for killing or even holding onto prey
because of the small size compared to other predatory dinosaurs.
Comparisons have even been drawn between Tyrannosaurus
and smaller
dromaeosaurid
dinosaurs like Velociraptor
as proof that Tyrannosaurus
could not hunt because it does not have the proportionately longer arms
that dromaeosaurid dinosaurs had.
Really
though it is not a case of arm length that determines if an animal is
an active predator or not, it is how an animal hunts. Dromaeosaurid
dinosaurs do have long arms, but they also have smaller heads and
comparatively weak bite forces when compared to larger carnivores.
Dromaeosaurid dinosaurs instead probably relied upon the enlarged
curved claws that were on their feet and it is generally accepted that
they used these claws to stab at their prey. In order to hit the
right weak spots like the necks of their victims, they would have had
an easier time of it if they reached up and held onto the prey animal
so that they moved with the animal as it struggled to escape,
meaning that they would only have to concentrate on moving their legs
to strike. In short, dromaeosaurids had longer arms because they
were necessary.
Tyrannosaurus
did not have specially adapted killing claws on its feet. It did
however have a large and massively powerful head that could rival any
other of the large theropod dinosaurs. This head was also carried at
a height that would have allowed for easy bites to the back and necks
of the larger dinosaurs of its time and location. All Tyrannosaurus
would have to do is come up alongside its prey and deliver a single
well placed bite to its spine to cripple if not kill its prey
instantly.
There
is a very good piece of evidence that supports the above hunting
strategy for Tyrannosaurus, and this comes from
the remains of an
Edmontosaurus specimen that shows signs of a Tyrannosaurus
attack.
Several of the vertebrae show a bite mark created by a Tyrannosaurus,
but more importantly the bones show that the wound healed. This
proves that the Edmontosaurus was alive when the Tyrannosaurus
bit into
it. Had it already been dead, the wound would not have healed,
although a few centimetres further down and the story would have been
very different.
Other
scavenger claims include the bone crushing bite and bone fragments
found in Tyrannosaurus coprolites. However,
many of the potential
prey species available to Tyrannosaurus were
armoured, and would
necessitate the bone crushing teeth. Also, the type of teeth in the
mouth of Tyrannosaurus would mean that flesh would
have to be ripped
from the carcass, and this process would undoubtedly pull bone free
whether Tyrannosaurus intended to or not.
The
olfactory senses of Tyrannosaurus were also highly
developed, giving
it an exceptional sense of smell. This indication could go either way
to support the scavenger or hunter theories for Tyrannosaurus
as a good
sense of smell is a vital part of any carnivorous animal.
At
the end of this, almost all predators that kill their own prey will
also actively scavenge carcasses as well. It just makes good sense as
scavenging requires far less calories than chasing and attacking prey,
and does not carry the risk of injury from struggling prey in the
process.
Tyrannosaurus
biology
The
actual non feeding lifestyle and biology of Tyrannosaurus
are also
subjects of interest. Because the back of the skull was wide in
proportion to the tip of the snout, Tyrannosaurus
had very good
'binocular vision'. This means that it had a good sense of depth
perception and knew exactly when another dinosaur was close enough to
strike.
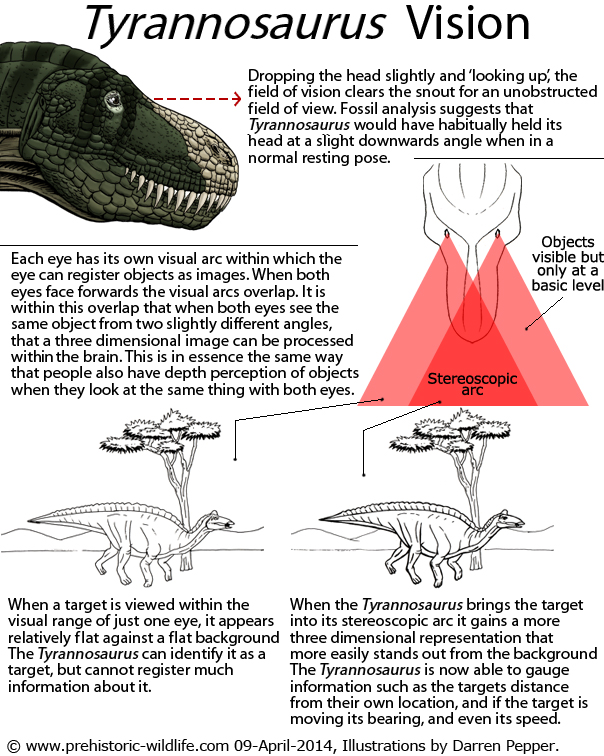
Returning
to the arms, although they were small, they were surprisingly
powerful. Biomechanical studies have suggested that the arms could
lift up to 200kg, enough to lift almost any human being off the
ground. The simple structure of the arms also indicates that they
were better suited to provide firm support, and may have been used by
Tyrannosaurus to grip another as they mated.
While
the arms do seem to have served some purpose in the life of
Tyrannosaurus, it has caused a lot of wonder as to
what would have
happened to them had the dinosaurs not gone extinct at the end of the
Cretaceous. Earlier members of the group like Eotyrannus
had
proportionately longer arms than Tyrannosaurus,
and the study of
group shows that as tyrannosaurids grew bigger, the arms got
shorter. Some think that given enough time, the arms may have
disappeared almost completely, like whales losing the hind limbs that
were present in their quadrupedal ancestors. Unfortunately, we will
probably never be certain of the answer.
The
jaws of Tyrannosaurus were tremendously powerful
with a bite force of
larger individuals being measured in several tons. The teeth were
primarily arranged into biting incisors at the front, with larger
canines to the size and rear of the mouth. The incisors that are on
the premaxilla form a 'D shaped' curve around the front, which is
notable as this gave Tyrannosaurus (and others of
its group) a 'U
shaped' snout. Other well-known carnivorous theropods such as
Allosaurus
were more 'V shaped'. The 'U'
shaped snout means
that Tyrannosaurus had more 'pick-up' every
time it took a bite
than other dinosaurs. The
teeth towards the sides of the mouth were the most suited to crushing,
and grow anywhere up to 30 centimetres long when including the
root. The teeth of the lower jaw (mandible) were smaller than
those above, and fitted inside the upper jaw so that when closed,
only the upper teeth would protrude out and downwards.
Tyrannosaurus
like so many of the other early discovered dinosaurs was once
considered to be a cold blooded and sluggish until warmed up in the sun
creature. Yet as a large theropod, Tyrannosaurus
can actually
present a case for a warm blooded metabolism through gigantothermy.
This is where the surface area of the animal is small when compared to
the volume and mass of the animal. A smaller surface area results in
a reduced area for heat to escape out of the body. This in turn
raises the base temperature of the animal beyond what it would be if it
were the same size but with a much thinner build. The
rapid growth curve exhibited in Tyrannosaurids as a
whole can also be
considered indicative of a warm blooded metabolism, as animals that
are cold blooded tend to have a more stable and constant rate of growth
until they are full grown.
It
has also been put forward that when very young, Tyrannosaurus
may
have had some feather growth as insulation on its body. This is based
upon the discovery that some of the basal and smaller tyrannosaurids
like Dilong
have been found with the impressions of protofeathers with
their remains. However what is known for fact is that as an adult,
Tyrannosaurus would have almost certainly had no
feathers at all,
with skin impressions of an adult Tyrannosaurus
revealing that it had
a bare pebble like texture to it.However, ever since the description of
Yutyrannus
in 2012, the notion that large tyrannosaurs may have had at
least a partial feather covering has now been seriously reconsidered.
Tyrannosaurus
remains can be divided into two groups of robust (larger and
heavier) and gracile (smaller and lighter) forms. For a long
time it was believed that these represented male and female specimens
with the larger ones presumably being female to allow for egg
development inside of the hips. Problems would eventually arise with
this thinking however.
It
was also thought that females had a reduced chevron in the first
caudal (tail) vertebra to allow for the passage of eggs.
However, comparison between remains show that first caudal vertebrae
with a fully developed chevron are also present in the robust (and
once presumed female) morphs. This means that the two morphs can no
longer be relied upon to tell the differences between male and female,
and are now instead considered to represent older individuals for the
robust morphs, younger for the gracile morphs. They may even be
representative of Tyrannosaurus populations hailing
from different
geographical areas.
Although
telling the difference between male and female Tyrannosaurus
is now
more problematic, one specimen can be confirmed as being absolutely
100% female. This is because of the presence of what is called
medullary tissue in the fossilised remains. In birds this tissue is a
kind of a calcium reserve that is used for the production of eggshells
during ovulation. Because it is used for this process, it obviously
only occurs naturally in female birds, and by extension, could only
exist in female Tyrannosaurus as they are the ones
that would have laid
the eggs, not the males. Remnants
of medullary tissue have also been found in some other dinosaurs, and
since its initial discovery in fossils, its presence has been
actively sought out in other specimens. The existing medullary tissue
found is taken as further evidence for the support for the theory that
birds evolved from dinosaurs.
An
exciting discovery about Tyrannosaurus biology came
about in 2005
when a femur was broken to facilitate transportation. Inside were
the remains of fibrous bone matrix tissue and blood vessels. It has
long been presumed that soft tissue is incompatible with the known
forms of fossilisation, but this has been seen as evidence that it is
on certain occasions possible to a certain extent. This discovery has
also helped lead to other fossils being tested for the presence of soft
tissue.
Further
study on soft tissue has led to analysis of collagen proteins from the
inner parts of the collagen coil, with results showing that they
closely match those of modern day chickens. Again this has reinforced
the theory that dinosaurs are indeed more closely related to birds.
Tyrannosaurus interaction
Although
there is some evidence to suggest pack hunting in some tyrannosaurs
like Albertosaurus
and Yutyrannus,
there is none present for Tyrannosaurus
itself.
Some Tyrannosaurus skulls bear marks that are
considered by some to
have been caused by the teeth of another Tyrannosaurus.
This would
bring in potential intraspecific biting that is seen in some carnivores
as signs of a dominance struggle. If true, this could present a
scenario where a Tyrannosaurus challenged another
in conflict over a
carcass, resulting in the bite marks to the face.
However,
a later theory is that these marks were not caused by another
Tyrannosaurus, but a parasite similar to one that
belongs to the
genus Trichomonas, a protist parasite that is
known to affect birds
today. These parasites cause growths in the soft tissue that can also
scar the bone underneath. Infection could have been caused quite
easily when a Tyrannosaurus drunk from water where Trichomonas
was
active, possibly left there by another infected dinosaur.
A
study undertaken in 2010 has brought forward the suggestion of
cannibalism in Tyrannosaurus. This comes from
what
appear to be strong
tooth marks to the legs and feet of a Tyrannosaurus
specimen. It is
thought that this individual was already dead when eaten because in a
living Tyrrannosaurus, the lower legs would have
been the furthest
points away from the mouth of another Tyrannosaurus,
making them very
hard to reach and not really viable targets when you consider that the
head and neck would be much easier to grab hold of. It is not certain
however if this Tyrannosaurus had been eaten by the
other that killed
it, or if it died from some other cause. Also these body parts have
some of the smallest amounts of muscle mass, further indicating the
Tyrannosaurus that was doing the eating had found a
carcass that had
already been picked clean of the choice pieces of flesh.
Tyrannosaurus
rex
or
Manospondylus gigas?
Technically,
Tyrannosaurus is not the original name for this
dinosaur. In
1892, Edward Drinker Cope named two incomplete vertebrae that he
thought were from a ceratopsian dinosaur. He named the new dinosaur
as Manospondylus gigas, which translates to
English as 'giant
porous vertebra'. Although lacking in completeness, the bones
were unknown and considered enough to warrant a genus name. What
needs to remembered is that this naming took place towards the end of
the 'Bone wars', a fierce rivalry between cope and Othniel
Charles Marsh that included who could name the most species. Today,
a naming for such scant material would be done only after very careful
consideration on the part of the researchers.
In
1905, Henry Fairfield Osborn named what was considered the first
specimen of Tyrannosaurus that was originally dug
up by Barnum Brown
in 1902. In a 1917 paper Osborn discussed many dinosaurs, but
also noted the similarity between the vertebra of Tyrannosaurus
and
Manospondylus. Despite these similarities being
pointed out,
nothing further happened until the turn of the following century.
In
the year 2000, the Black Hills Institute of Geological Research
found what is believed to be the original fossil site for Manospondylus
gigas. There, the dig crew recovered further Tyrannosaurus
remains
confirming that the Manospondylus remains did
indeed belong to
Tyrannosaurus. This
presented immediate upset not just in the paleontological community but
in news reports around the world. Typically under ICZN rules, the
first name has priority of any subsequent names that are given to the
same genus. In this scenario, Tyrannosaurus
would only exist as a
synonym, and all fossils and future references to Tyrannosaurus
would
have to say Manospondylus instead.
However
a revision to ICZN rules that happened a little earlier in 2000 may
actually preserve the name Tyrannosaurus. The
simplest version of
these revisions come across as any junior synonym (in this case
Tyrannosaurus) that has been used as the accepted
name for a species
instead of the senior name (Manospondylus) that
was granted before
1899, may be used as the official name for that taxon.
This
means that Tyrannosaurus is likely going to remain
known as
Tyrannosaurus because of so many studies published
by so many
palaeontologists referring to it as Tyrannosaurus.
In contrast,
Manospondylus is virtually unknown if you don't
count the very scant references to
its similarity with Tyrannosaurus, and as such
will become treated in
a similar manner to any other synonym to Tyrannosaurus.
Tyrannosaurus,
Tarbosaurus and
other suggested synonyms
Scant
remains of tyrannosaurs such as teeth and the occasional post cranial
bone were being found during the late nineteenth century, but it was
not until 1905 when the first Tyrannosaurus
specimen was described
that people had an idea what they were dealing with. This is why
Tyrannosaurus has been used to name the groups
Tyrannosauoirdea,
Tyrannosauridae and Tyrannosaurinae, there was enough left of this
genus to identify remains of other similar dinosaurs as tyrannosaurs.
Before the term tyrannosaur came into use however, tyrannosaurs
were known as deinodonts after the genus Deinodon
(‘terrible
tooth’), but like with many of the genera established before the
description of Tyrannosaurus, it was based mostly
on teeth, which
means that it would be incredibly difficult to attribute skeletal
remains to the genus with certainty.
In
1955 an expedition in Mongolia recovered the remains of a huge
tyrannosaur that bore a superficial resemblance to Tyrannosaurus,
and
even today, if you only took a quick glance at the side view of these
fossils you would be hard pressesd to spot a clear difference. The
palaeontologist Evgeny Maleev described these remains as a new species
of Tyrannosaurus, T. bataar,
though almost immediately there was
some concern about whether this was the right interpretation. Ten
years later in 1965 Tyrannosaurus bataar was
renamed Tarbosaurus
bataar, and from then on there has been a nearly
constant debate over
how to keep the fossils named, as a species of Tyrannosaurus
or the
new genus Tarbosaurus.
Most
studies concerning these two tyrannosaurs have centred on finding
differences and similarities between them, but what defines enough
differences to registers a new species or a new genus can vary greatly
amongst researchers. While the skulls of Tyrannosaurus
and
Tarbosaurus are quite similar when seen from the
side, when viewed
from the front or above it becomes immediately clear that the skull of
Tarbosaurus is far narrower than the skull of Tyrannosaurus.
Further
analysis has also shown that the way that stresses are transmitted
through the skull is very different in Tarbosaurus.
Tarbosaurus
was once widely accepted as the sister taxon to Tyrannosaurus,
but
bringing other genera into the equation this is no longer the case.
Another tyrannosaur from Mongolia that was once considered to be a
juvenile Tarbosaurus is Alioramus.
Studies and new fossils of
Alioramus now not only confirm that it is indeed a
distinct genus,
but that it is actually closer to Tarbosaurus than
what Tyrannosaurus
is, something that suggests no immediate familial link between
Tarbosaurus and Tyrannosaurus
other than they were both tyrannosaurs.
The discovery of the genus Lythronax in North America has also
indicated that the tyrannosaurids were present their long before the
appearance of either Tyrannosaurus in North America
or Tarbosaurus in
Asia. Today, most palaeontologists agree that it is a better idea
to keep Tyrannosaurus and Tarbosaurus
as separate genera, especially
as more studies and new remains point more to this conclusion.
For
a long time now there has been a ‘big four’ tyrannosaur genera in
North America which would have lived in upper Laramidia at the end of
the Cretaceous. These are Tyrannosaurus, Daspletosaurus,
Albertosaurus and Gorgosaurus,
the latter three of which
occasionally get treated as synonyms. Albertosaurus
and Gorgosaurus
are quite different to Tyrannosaurus in that they
are more lightly
built. Gorgosaurus is frequently synonymised with
Albertosaurus too,
yet at this time Gorgosaurus is still regarded as
valid by many.
Albertosaurus is and by extension Gorgosaurus
are perhaps the least
likely to ever be synonymised to Tyrannosaurus due
to their obviously
lighter builds. Daspletosaurus is more robust
like Tyrannosaurus,
but so far known from slightly earlier in the Cretaceous. Some
continue to treat Daspletosaurus as a synonym to Tyrannosaurus,
though most choose to keep the genus separate. In addition to this,
some people consider Daspletosaurus to be the
genus that is directly
ancestral to the Tyrannosaurus genus.
Further reading
- Tyrannosaurus and other Cretaceous carnivorous
dinosaurs - Bulletin
of the AMNH - Henry Fairfield Osborn - 1905.
- Tyrannosaurus, Upper Cretaceous carnivorous
dinosaur - Bulletin of
the AMNH (New York City: American Museum of Natural History) 22 (16):
281–296 - Henry Fairfield Osborn & Barnum Brown - 1906.
- Skeletal adaptations of Ornitholestes, Struthiomimus,
Tyrannosaurus".
Bulletin of the American Museum of Natural History (New York City:
American Museum of Natural History) 35 (43): 733–771 - Henry Fairfield
Osborn - 1917.
- [Gigantic carnivorous dinosaurs of Mongolia], E. A. Maleev
(translated by F. J. Alcock) - Doklady Akademii Nauk SSSR - 1955.
- Growth changes in Asian dinosaurs and some problems of their taxonomy
- Paleontological Journal 3: 95–109. - A. K. Rozhdestvensky - 1965.
- Stance and gait in the flesh-eating Tyrannosaurus
- Biological
Journal of the Linnean Society 2 (2): 119–123 B. H. Newman - 1970.
- Tiny dinosaurs: Are they fully grown? - Journal of Vertebrate
Paleontology 3 (4): 200–209. - G. Callison & H. M. Quimby -
1984.
- Long bone circumference and weight in mammals, birds and dinosaurs -
Journal of Zoology 207 (1): 53–61 - J. F. Anderson, A. J. Hall-Martin
& D. A. Russel - 1985.
- Nanotyrannus, a new genus of pygmy tyrannosaur,
from the latest
Cretaceous of Montana - Hunteria 1 (5): 1–30 - R. T. Bakker, M.
Williams & P. J. Currie - 1988.
- Variation in Tyrannosaurus rex, Kenneth Carpenter
- In Kenneth
Carpenter and Philip J. Currie. Dinosaur Systematics: Approaches and
Perspectives. Cambridge: Cambridge University Press. pp. 141–145 - 1992.
- The serrated teeth of tyrannosaurid dinosaurs, and biting structures
in other animals - Paleobiology18 (2): 161–183 - Abler William - 1992.
- Steak knives, beady eyes, and tiny little arms (a portrait of
Tyrannosaurus as a scavenger) - The Paleontological
Society Special
Publication 7: 157–164 - J. R. Horner - 1994.
- The Phylogenetic Position of the Tyrannosauridae: Implications for
Theropod Systematics - Journal of Palaeontology 68 (5): 1100–1117 -
Thomas R. Holtz - 1994.
- A track of the giant theropod dinosaur Tyrannosaurus
from close to
the Cretaceous/Tertiary boundary, northern New Mexico - Ichnos 3 (3):
213–218. - M. G. Lockley & A. P. Hunt - 1994.
- Thermophysiology of Tyrannosaurus rex: Evidence
from Oxygen Isotopes
- Science (New York City) 265 (5169): 222–224 - Reese E. Barrick
& William J. Showers - 1994.
- Body mass, bone "strength indicator", and cursorial potential of
Tyrannosaurus rex - Journal of Vertebrate
Paleontology 15 (4): 713–725
- J. O. Farlow, M. B. Smith & J. M. Robinson - 1995.
- Bite-force estimation for Tyrannosaurus rex from
tooth-marked bones -
Nature 382 - G. M. Erickson, S. D. Van Kirk, J. Su, M. E. Levenston, W.
E. Caler & D. R. Carter - 1996.
- Head-biting behavior in theropod dinosaurs: paleopathological
evidence - Gaia (15): 167–184 - Darren H. Tanke & Phillip J.
Currie - 1998.
- Strength indicator values of theropod long bones, with comments on
limb proportions and cursorial potential - Gaia 15: 241–255 - P.
Christiansen - 1999.
- Estimating the masses and centers of mass of extinct animals by 3-D
mathematical slicing - Paleobiology 25 (1): 88–106 - Donald M.
Henderson - 1999.
- Craniofacial ontogeny in Tyrannosauridae (Dinosauria, Theropoda) -
Journal of Vertebrate Paleontology 19 (3): 497–520. - T. D. Carr - 1999.
- Thermophysiology and biology of Giganotosaurus:
comparison with
Tyrannosaurus - Palaeontologia Electronica 2 - Reese
E. Barrick
& William J. Showers - 1999.
- Limb design, function and running performance in ostrich-mimics and
tyrannosaurs - Gaia 15: 257–270 - Gregory S. Paul - 2000.
- A new method to calculate allometric length–mass relationships of
dinosaurs - Journal of Vertebrate Paleontology 21 (1): 51–60 - Frank
Seebacher - 2001.
- Forelimb Osteology and Biomechanics of Tyrannosaurus rex,
Kenneth
Carpenter & Matt Smith - In Darren Tanke & Kenneth
Carpenter, Mesozoic vertebrate life. Bloomington: Indiana University
Press. pp. 90–116 - 2001.
- Tyrannosaurus was not a fast runner - Nature 415
(6875): 1018–21 - J.
R. Hutchinson & M. Garcia - 2002.
- Maximum bite force and prey size of Tyrannosaurus rex
and their
relationships to the inference of feeding behavior - Historical
Biology: A Journal of Paleobiology 16 (1): 1–12 - Mason B. Meers - 2003.
- Diagenetic effects on the oxygen isotope composition of bones of
dinosaurs and other vertebrates recovered from terrestrial and marine
sediments - Journal of the Geological Society 160 (6): 895 - Clive
Trueman, Carolyn Chenery, David A. Eberth & Baruch Spiro - 2003.
- Giant theropod dinosaurs from Asia and North America: Skulls of
Tarbosaurus bataar and Tyrannosaurus rex
compared - Acta
Palaeontologica Polonica 48 (2): 161–190 - J�rn H. Hurum &
Karol Sabath - 2003.
Could Tyrannosaurus rex have been a scavenger
rather than a predator?
An energetics approach - Proceedings. Biological sciences the Royal
Society 270 (1516): 731–3 - G. D. Ruxton & D. C. Houston - 2003.
- Osteology of Tyrannosaurus rex: insights from a
nearly complete
skeleton and high-resolution computed tomographic analysis of the skull
- Society of Vertebrate Paleontology Memoirs 7: 1–138 - C. R. Brochu -
2003.
- Cranial anatomy of tyrannosaurid dinosaurs from the Late Cretaceous
of Alberta, Canada - Acta Palaeontologica Polonica 42 (2): 191–226. -
Phillip J. Currie - 2003.
- Skull structure and evolution in tyrannosaurid dinosaurs - Acta
Palaeontologica Polonica 48 (2): 227–234. - Philip J. Currie, J�rn H.
Hurum & Karol Sabath - 2003.
- Diversity of late Maastrichtian Tyrannosauridae (Dinosauria:
Theropoda) from western North America - Zoological Journal of the
Linnean Society 142 (4): 479–523 - T. D. Carr & T. E.
Williamson - 2004.
- Obligate vertebrate scavengers must be large soaring fliers - Journal
of Theoretical Biology 28 (3): 431–436 - Graeme D. Ruxton &
David C. Houson - 2004.
- Gigantism and comparative life-history parameters of tyrannosaurid
dinosaurs - Nature 430 (7001): 772–775 - Gregory M. Erickson, Peter
J.Makovicky, Philip J. Currie, Mark A. Norell, Scott A. Yerby &
Christopher A. Brochu - 2004.
- Mass prediction in theropod dinosaurs - Historical Biology 16 (2–4):
85–92 - Per Christiansen & Richard A. Fari�a - 2004.
- Age and growth dynamics of Tyrannosaurus rex -
Proceedings.
Biological sciences / the Royal Society 271 (1551) - J. R. Horner
& K. Padian - 2004.
- Soft-tissue vessels and cellular preservation in Tyrannosaurus
rex -
Science 307 (5717): 1952–5 - Mary H. Schweitzer, Jennifer L. Wittmeyer,
John R. Horner & Jan K. Toporski - 2005.
- Gender-specific reproductive tissue in ratites and Tyrannosaurus
rex
- Science 308 (5727): 1456–60 - M. H. Schweitzer, J. L. Wittmeyer
& J. R. Horner - 2005.
- In Celebration of 100 years of Tyrannosaurus rex:
Manospondylus
gigas, Ornithomimus grandis, and Dynamosaurus
imperiosus, the Earliest
Discoveries of Tyrannosaurus Rex in the West -
Abstracts with Programs.
2005 Salt Lake City Annual Meeting 37 (7). Geological Society of
America. p. 406 - Brent H. Breithaupt, Elizabeth H. Southwell &
Neffra A. Matthews - 2005.
- Heterodonty in Tyrannosaurus rex: implications
for the taxonomic and
systematic utility of theropod dentitions - Journal of Vertebrate
Paleontology 25 (4): 865–887 - J. B. Smith - 2005.
- Binocular vision in theropod dinosaurs - Journal of Vertebrate
Paleontology 26 (2) - Kent A. Stevens - 2006.
- Fused and vaulted nasals of tyrannosaurid dinosaurs: Implications for
cranial strength and feeding mechanics - Acta Palaeontologica Polonica
51 (3): 435–454. - Eric Snively, Donald M. Henderson & Doug S.
Phillips - 2006.
- Dynamosaurus imperiosus and the earliest discoveries of Tyrannosaurus
rex in Wyoming and the West - New Mexico Museum of Natural
History and
Science Bulletin 35: 257–258 - B. H. Breithaup, E. H. Southwell
& N. A. Matthews - 2006.
- Tyrannosaur life tables: an example of nonavian dinosaur population
biology - Science 313 (5784): 213–7 - G. M. Erickson, P. J. Currie, B.
D. Inouye & A. A. Winn - 2006.
- A 3D interactive method for estimating body segmental parameters in
animals: application to the turning and running performance of
Tyrannosaurus rex - Journal of Theoretical Biology
246 (4): 660–80 -
John R. Hutchinson, Victor Ng-Thow-Hing & Frank C. Anderson -
2007.
- Estimating dinosaur maximum running speeds using evolutionary
robotics - Proc. R. Soc. B (The Royal Society) 274 (1626): 2711–6 - W.
I. Sellers & P. L. Manning - 2007.
- Craniocervical feeding dynamics of Tyrannosaurus rex
- Paleobiology
33(4): 610-638 - Eric Snively & Anthony P. Russel - 2007.
- Tyrannosaurus rex, the Tyrant King (Life of the
Past). Bloomington:
Indiana University Press. ISBN 0-253-35087-5, Kenneth Carpenter
& Peter E. Larson - 2008.
- Sexual maturity in growing dinosaurs does not fit reptilian growth
models - Proceedings of the National Academy of Sciences 105 (2):
582–587 - Andrew H. Lee & Sarah Werning - 2008.
- Looking again at the forelimb of Tyrannosaurus rex".
In Carpenter,
Kenneth; and Larson, Peter E. (editors), Tyrannosaurus rex,
the Tyrant
King (Life of the Past). - Bloomington: Indiana University Press -
Christine Lipkin, & Kenneth Carpenter - 2008.
- Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms - In
Stepanova, Anna. PLoS ONE 3 (7): e2808 - Thomas G. Kaye, Gary Gaugler
& Zbigniew Sawlowicz - 2008.
- New insights into the brain, braincase, and ear region of
tyrannosaurs (Dinosauria, Theropoda), with implications for sensory
organization and behavior. - The Anatomical Record 292(9) 1266-1296 -
Lawrence M. Witmer & Ryan C. Ridgely - 2009.
- Common Avian Infection Plagued the Tyrant Dinosaurs - In Hansen,
Dennis Marinus. PLoS ONE 4 (9): e728, - E. D. S. Wolff, S. W.
Salisbury, J. R. Horner & D. J. Varricchi - 2009.
- Face Biting On A Juvenile Tyrannosaurid And Behavioral Implications -
Palaios 24 (11): 780–784 - Joseph E. Peterson, Michael D. Henderson
& Reed P. Sherer & Christopher P. Vittore - 2009.
- Cannibalism in Tyrannosaurus rex - Public Library
of Science. - N. R.
Longrich, J. R. Horner, G. M. Erickson & P. J Currie - 2010.
- The Tail of Tyrannosaurus: Reassessing the Size
and Locomotive
Importance of the M. caudofemoralis in Non-Avian Theropods - The
Anatomical Record: Advances in Integrative Anatomy and Evolutionary
Biology 294 (1): 119–131 - Scott W. Persons & Phillip J. Currie
- 2010/2011.
- A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus
rex with Implications for Locomotion, Ontogeny, and Growth -
PLoS ONE 6
(10): e26037 - J. R. Hutchinson, K. T. Bates, J. Molnar, V. Allen, P.
J. Makovicky - 2011.
- Tyrannosaurs were power-walkers - Nature - Matt Kaplan - 2011.
- Dinosaur Census Reveals Abundant Tyrannosaurus
and Rare Ontogenetic
Stages in the Upper Cretaceous Hell Creek Formation (Maastrichtian),
Montana, USA - PLoS ONE 6 (2): e16574 - John R. Horner, Mark B. Goodwin
& Nathan Myhrvold - 2011.
- Estimating maximum bite performance in Tyrannosaurus rex
using
multi-body dynamics - Biological Letters - K. T. Bates & P. L.
Falkingham - 2012.
- Tyrant Dinosaur Evolution Tracks the Rise and Fall of Late Cretaceous
Oceans - In Evans, David C. PLoS ONE 8 (11) - M. A. Loewen, R. B.
Irmis, J. J. W. Sertich, P. J. Currie & S. D. Sampson - 2013.
- The Posture of Tyrannosaurus rex: Why Do Student
Views Lag Behind the
Science? - Journal of Geoscience Education 61: 145 - R. M. Ross, D.
Duggan-Haas & W. D. Allmon - 2013.
- Physical evidence of predatory behavior in Tyrannosaurus rex.
-
Proceedings of the National Academy of Sciences - Robert A. DePalma II,
David A. Burnham, Larry D. Martin, Bruce M. Rothschild & Peter
L. Larson - 2013.
- Skin pathology in the Cretaceous: Evidence for probable failed
predation in a dinosaur - Cretaceous Research , 42(0),44-47 - Bruce M.
Rothschild & Robert DePalma - 2013.
- Chemistry supports the identification of gender-specific reproductive
tissue in Tyrannosaurus rex. - Scientific Reports.
6 (23099). - M. H.
Schweitzer, W. Zheng, L. Zanno, S. Werning & T. Sugiyama - 2016.
- The biomechanics behind extreme osteophagy in Tyrannosaurus
rex. -
Scientific Reports. 7 (1): 2012. - P. M. Gignac & G. M.
Erickson - 2017.
- Tyrannosauroid integument reveals conflicting patterns of gigantism
and feather evolution. - Biology Letters. 13 (6): 20170092. - P. R,
Bell, N. E. Campione, W. S. Persons IV, P. J. Currie, P. L. Larson, D.
H. Tanke & R. T. Bakker - 2017.
- Investigating the running abilities of Tyrannosaurus rex
using
stress-constrained multibody dynamic analysis. - PeerJ. 5: e3420. - W.
I. Sellers, S. B. Pond, C. A. Brassey, P. L. Manning & K. T.
Bates - 2017.
- Lower rotational inertia and larger leg muscles indicate more rapid
turns in tyrannosaurids than in other large theropods". PeerJ. 7:
e6432. - Eric Snively, Haley O’Brien, Donald M. Henderson, Heinrich
Mallison, Lara A. Surring, Michael E. Burns, Thomas R. Holtz, Jr,
Anthony P. Russell, Lawrence M. Witmer, Philip J. Currie, Scott A.
Hartman & John R. Cotton - 2019.
- Feeding traces attributable to juvenile Tyrannosaurus rex
offer
insight into ontogenetic dietary trends. - PeerJ. 7: e6573. - Joseph E.
Peterson & Karsen N. Daus - 2019.
- An Older and Exceptionally Large Adult Specimen of Tyrannosaurus
rex.
- The Anatomical Record. 303 (4): 656–672. - S. W. Persons, P. J.
Currie & G. M. Erickson - 2020.
- Growing up Tyrannosaurus rex: Osteohistology
refutes the pygmy
"Nanotyrannus" and supports ontogenetic niche
partitioning in juvenile
Tyrannosaurus. - Science Advances. 6 (1). - Holly N.
Woodward, Katie
Tremaine, Scott A. Williams, Lindsay E. Zanno, John R. Horner &
Nathan Myhrvold - 2020.
- Tracks in the upper cretaceous of the raton basin possibly show
tyrannosaurid rising from a prone position. - New Mexico Museum of
Natural History and Science: 29–37. - T. Canner, T. Molkestad &
S. G. Lucas - 2021.
Random favourites
 |
 |
 |
 |