































Archaeopteryx
a.k.a. Urvogel
Name:
Archaeopteryx
(Ancient wing). Archaeopteryx is sometimes
called Urvogel which is German for ‘first bird’, but this is not an
alternate binomial name.
Phonetic: Ar-kay-op-teh-rix.
Named By: Christian Erich Hermann von Meyer -
1861.
Synonyms: Archaeornis, Griphosaurus,
Griphornis, Jurapteryx, Pterodactylus crassipes, Rhamphorhynchus
crassipes, Scaphognathus crassipes. Possibly also Wellnhoferia.
Classification: Chordata, Aves,
Archaeopterygidae.
Species: A. lithographica
(type). A. albersdoerferi. Additional species
previously established include Archaeopteryx
bavarica, Archaeopteryx owenii, Archaeopteryx macrura,
Archaeopteryx recurva and Archaeopteryx siemensii,
but there validity is not in agreement by all. See main text for
details.
Diet: Carnivore/Insectivore.
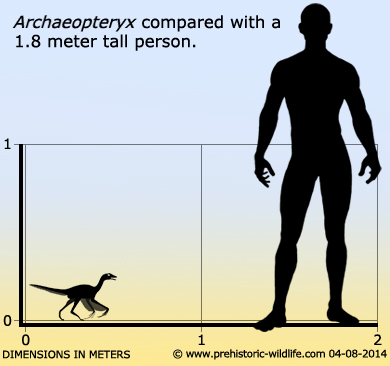
Size: Up to about 50 centimetres long.
Known locations: Various locations in Germany.
Time period: Tithonian of the Jurassic.
Fossil representation: Many specimens, some almost
complete and preserved flat on a slab, some just of feathers.
Archaeopteryx
- The
controversy and the facts
Very
few genera have shaken both the worlds of palaeontology and
evolutionary theory as much as Archaeopteryx. The
story begins in the
limestone quarries near Solnhofen, Germany. The limestone here is
the result of geologic processes changing deposits of aquatic sediment
all the way back to the late Jurassic. Here the fossils of all kinds
of creatures from ammonites to fish to even pterosaurs
have been
found, but it was one fossil in particular that caught the eye of
Christian von Meyer; the fossil of a feather.
The
fossil of a feather by itself might not sound interesting, but you
need to realise that this discovery took place in 1861 when the
scientific field of palaeontology was still in its infancy. The idea
of extinct animals that roamed the land before man had only been
properly realised in the early nineteenth century. The first
pterosaur Pterodactylus
had been named in 1809, the first dinosaur
Megalosaurus
in 1824 and Charles Darwin’s On the Origin of Species
had only been published a couple of years earlier in 1859. The
truly radical thing about the discovery of this single feather
impression was that it came from a Jurassic aged deposit, a fact that
shook the then and relatively new theory that birds did not appear
until the Eocene after the dinosaurs had disappeared.
Christian
von Meyer named a new genus in 1861, Archaeopteryx
lithographica,
with the feather as holotype, but as you might expect naming a new
genus of bird by only a feather means that it is hard to accurately
reconstruct the animal. But in the same year a new fossil was found
at Solnhofen, and this time it was a partial skeleton. Now known as
the London Specimen (or BMNH 37001 for its catalogue number),
this fossil was first given to a physician named Karl H�berlein who
went on to sell it to the National History Museum of London for
�700, a sum of money that back then was worth far more than it is
today. The London Specimen was described 1863 by Richard Owen,
who was one of the chief naturalists in Britain at the time. Owen
recognised that the skeleton probably belonged to the same kind of
creature as the feather, but also recognised the difficulty in
assigning skeletal remains to only a feather. So with this in mind
Owen created a second species, A. macrura,
though this would be
the start of further controversy discussed more fully in the next
section.
Missing
only the head and neck, the London Specimen gave the first true
glimpse at what Archaeopteryx looked like.
However, the skeletal
remains were not those of a bird that we would recognise today. The
most obvious difference was the tail, long and lizard-like, but
there were also two free claws on each wing. This came as a bit of a
shock to those who were expecting Archaeopteryx to
actually look more
like a bird, but for evolutionists the find was a dream come true.
Evolutionary theory back in the nineteenth century was still a very
new idea, one that many naturalists considered to be complete
nonsense. To the evolutionists however, the London Specimen was
proof that the birds did not just magically appear and that their
origins could be found within the reptiles.
Around
1874/1875 a new specimen of Archaeopteryx was
found by a farmer name
Jakob Niemeyer. Niemeyer sold this to an inn keeper named Johann D�rr
in 1876 before it was later sold to Ernst Otto H�berlein. The
specimen was then later sold at auction to Ernst Werner von Siemens and
donated to the Humboldt Museum f�r Naturkunde. Today known as the
Berlin Specimen (HMN 1880), this was the first specimen to
include the head, and again things were not as expected. The skull
seen here was also more reptile-like and the jaws still had conical
teeth within them.
After
this time many other specimens began to be discovered, and these
discoveries have continued into the twenty-first century. The Maxberg
Specimen (S5), an almost complete skeleton lacking only the head
and tail was discovered in 1956 and described in 1959. In
1970 it was discovered the Christian von Meyer, the man who named
the first feather in 1861, had actually handled and named the first
known specimen of Archaeopteryx in 1857, but
had mistakenly named
it as the pterosaurs Rhamphorhynchus
crassipes and Pterodactylus
crassipes Described from partial post cranial bones, this individual
is now known as both the Haarlem Specimen and the Teyler Specimen
(TM 6428).
The
smallest individual was found in 1951 and named in 1974 is the
Eichst�tt Specimen (JM 2257). Originally thought to be a
specimen of the dinosaur Compsognathus,
the Solnhofen Specimen
(BSP 1999) was named in 1988. The Munich Specimen (S6) was
described in 1993 and represents an almost complete specimen
missing only the anterior (front) portion of its face. The
Daiting Specimen was discovered in 1990 and although fragmentary,
is known from slightly younger deposits than the Solnhofen specimens.
The B�rgermeister-M�ller Specimen is of a single wing and was
discovered in 2000 and described in 2004. After this the
Thermopolis Specimen was described in 2005, and is known by an
almost complete skeleton that more clearly displays theropod
dinosaurian features. An additional specimen of what is thought to be
Archaeopteryx was discovered in 2011, but at the
time of writing
this has not been made available for scientific description.
While
all of these new specimens were being found, some palaeontologists
were working upon piecing together the wider evolutionary link between
birds and reptiles, although during this time the link was narrowed
down to the maniraptorian theropod dinosaurs. Other primitive birds
were being found but to further confuse things, the existence of some
dinosaurs that had feathers covering their bodies became established
fact. The exact line between bird-like dinosaurs and dinosaur-like
birds is blurry to say the least, and opinions on where to draw it
can vary greatly amongst different palaeontologists. In the past,
Archaeopteryx has been considered to be both a
dinosaur and a bird,
but most thinking by the early twenty-first century places it as a
bird. In a wider sense Archaeopteryx is more
universally treated as a
transitional form that shows a stage in the evolutionary change between
dinosaurs and birds.
Archaeopteryx
has been accused of not being a transitional form and even not being
real upon numerous occasions. These accusations however have always
come from people who do not know what they are talking about with very
little to no understanding of fossilisation processes as well as wider
discoveries in other areas. A lot of people who have accused
Archaeopteryx of being fake are also not even aware
that the genus is
known by more than one specimen, or at least choose not to tell
people that they are trying to convince. Most of these claims go back
into the nineteenth and twentieth centuries, but the rise of personal
blogging during the early twenty-first century has meant that many of
these old and since long discredited theories are now being put forward
as ‘fact’ again.
Great
care should be taken not to confuse Archaeopteryx
with
‘Archaeoraptor’,
the name of an actual fake composite fossil that
came to light in 1999.
Which species of Archaeopteryx
are valid?
This
is a good question but one that has no universally accepted answer at
this time. There have been many names and species put forward to
represent specimens of Archaeopteryx, but many of
these to begin with
were either unnecessary or contained spelling/grammatical errors that
had to be later altered.
The
first complication comes from a spat between Christian von Meyer and
another German palaeontologist named Johann Andreas Wagner who proposed
the alternative Griphosaurus in 1862. Another
1862 alternative
put forward by Woodward was Griphornis, so now
there were three names
for one animal. The Pterodactylus crassipes
confusion was still not
known at the time, but if it were it would have had no more bearing
than the type species being changed to A. crassipes
because
Archaeopteryx obviously was not a pterosaur.
Things
began to be cleared up a little in 1951 when Gavin de Beer treated
the London Specimen, previously named A. macrura,
as the holotype
of the genus because this was at least a partial skeleton rather than
just a feather. The type species of A. lithographica
had already
been established for the original feather description, so A.
macrura became a synonym to A. lithographica.
This was backed up
in 1960 by another named Swinton. The ICZN, the international
scientific body that governs the naming of animals not only agreed with
de Beer, they also suppressed the earlier alternative names for
Archaeopteryx so that they were then known as
synonyms for the clarity
of later generations, listed but no longer used.
This
meant that the type species of Archaeopteryx was
now represented by the
original feather of 1861 and the first known skeletal remains of
1863, but another problem was about to raise its head. Closer
study of the feather with the later referred remains resulted in the
observation that the original feather was not a perfect match, and
may have actually come from a different bird, or perhaps even a
feathered theropod dinosaur. Apart from possibly being different,
in modern palaeontology a single feather would be considered to be too
indeterminate to describe a new genus, and beginning in 2007 this
was pointed out by two groups of scientists that petitioned the ICZN to
recognise the London Specimen remains instead of the feather.
Although the London Specimen remains had been proposed in 1951 by
de Beer, and added to the type species, they could never become the
holotype because the feather was already listed. Instead in 2011
the London Specimen became the neotype, the term for a fossil
specimen that is added to a genus after the holotype, but becomes the
set of remains that future discoveries are referred to instead of the
holotype. Today the original feather is considered just that, a
feather of indeterminate origin, probably not Archaeopteryx.
The
type species of Archaeopteryx with the London
Specimen designated as
neotype is the only species that has been universally accepted as
valid. The other specimens of Archaeopteryx have
all been credited as
different species due to differences in things such as form and size,
but a few of these species have been questioned. Some of the
specimens of Archaeopteryx can form to create small
groups. The
London (A. lithographica), Berlin (A.
siemensii) and
Haarlem specimens are fairly similar to one another. But the Munich
(A. bavarica), Eichst�tt (A.
recurva), Thermopolis
(A. siemensii) and Solnhofen specimens are
noted as having more
slender snouts, forward pointing teeth, possible presence of
sternums and being either smaller of larger than the London, Berlin
and Haarlem specimens. A. recurva was also
originally described as
a different genus, Jurapteryx, while the
Solhofen specimen is
particularly large and was renamed as a new genus, Wellnhoferia,
by
Andrzej Elzanowski in 2001, though this has had a mixed reception
with a 2003 study by Senter and Robins supporting this, but a
2007 study by Mayr et al. finding Wellnhoferia to still be
specimen of Archaeopteryx lithographica.
One
argument that may one day unify all of the specimens under
Archaeopteryx is that they simply represent
different growth stages
throughout the lifespan of Archaeopteryx. The
precedent for this has
actually already been set in other genera’s of prehistoric creatures,
the pterosaur genera Pterodactylus and Rhamphorhynchus
being two good
examples. Both of these genera were once each known by multiple
species, and while some of these species were later named as new
genera, many of them were realised to be just juveniles and subadults
that had differing physical proportions to adults. In time, more
specimens of Archaeopteryx will likely be
discovered given that so many
are already known, and these together may conclusively prove one way
or another which theory or indeed a measure of both is more correct.
Could Archaeopteryx
fly?
By
the early twenty-first century Archaeopteryx is
most widely considered
to be a bird, but one with many dinosaur-like characteristics.
Quite surprisingly, Archaeopteryx has been
considered to have been
similar to a modern bird called the Hoatzin (Opisthocomus
hoazin)
which is native to upper South America. Although not a perfect
match, juvenile Hoatzin possess claws that enable them to climb
branches until their wings are strong and developed enough to enable
flight.
Being
popularly defined as a bird. the obvious question would be could
Archaeopteryx fly? It would seem that the answer
depends on whether
you mean powered flight or just gliding, but first we need to
consider what is necessary to enable flight in the first place. First
and foremost would be the feathers since birds don’t rely upon
extended flaps of skin or membranes like other flying/gliding
vertebrates. Birds require special kinds of feathers which as you
might guess are called ‘flight feathers’, and these are found on
the rear edges of the wings for primary lift, and the tail for
steering. Aside from lift these feathers can also be angled by wing
and tail movements to alter things like drag to control flying speed in
a similar fashion to the flaps on the wing of an aeroplane (in fact
engineers copied this from watching birds).
A
flight feather is generally much longer than it is wide and is
reinforced by a rigid spine that runs down the length of the feather so
that it keeps straight and does not bend with the up and down movements
of the wing. These kinds of feathers are like those on the wings of
Archaeopteryx, which means that the genus has at
least one of the
things necessary for flight. The flight feathers on Archaeopteryx
though are not exactly like the feathers of modern birds because they
lack things called barbicles. Barbicles are like tiny hooks on the
fronds of the feather that hook around the fronds of the feather they
are next to, the barbicles of that feather then hook onto the next
and so on. Barbicles are simple features but they allow all of the
feathers on the wing to act as one large complete unit rather than lots
of smaller individual ones. This reinforces the wing, making the
feathers not only stronger but capable of providing more lift. The
fact that Archaeopteryx lacked these barbicles does
not mean that they
could not fly, just that Archaeopteryx were not
yet evolved into
modern forms, something we already knew. As time went on and
barbicles began to appear, they became more standardised, but the
principal here is that they had to evolve somewhere, and it seems
that it was not with Archaeopteryx, but possibly
a later and more
advanced form.
The
next thing to consider is was Archaeopteryx capable
of producing a
‘flight stroke’? A flight stroke is a movement of the wings that
allows for the production of lift to get the body into the air, and
then repeating that stroke or a variation of to stay in the air. The
key to producing a flight stroke is to have flexibility, ideally to
be able to move the wings (which remember are really just modified
arms) above the back for an extended down stroke which can produce a
greater amount of lift. The analysis of Archaeopteryx
skeletons
however is not promising. In modern birds the shoulder is aligned
dorsally (to the spine) whereas in Archaeopteryx
the shoulder has a
sideways alignment. This is a throwback to more dinosaurian
ancestors, but it terms of flight it means that the wings of
Archaeopteryx did not have the same range of motion
as modern birds.
This means that at best if Archaeopteryx could
fly, it could not fly
as well as modern birds.
A
reduced range of motion does not mean that Archaeopteryx
could not take
to the air. By holding the arms out to the sides it is conceivable
that Archaeopteryx could be gliders since the
developed flight feathers
of the wings would have still acted to reduce the rate of descent.
The wings of Archaeopteryx were fairly short and
rounded on the ends,
similar to birds that live in woodland environments today. Assuming
Archaeopteryx lived in these environments, and it
seems quite
possible, then they may have climbed trees with the claws on their
wings for either food or shelter, and then jumped and glided to the
next tree along. But so far we have only looked at two areas that are
necessary for flight, and from here we need to go from beyond just
wings and feathers.
Powered
flight by flapping wings requires the development of muscles capable of
maintaining the same repetitive movement for a long time, and these
muscles in turn require skeletal attachments to secure them in place.
The muscles for flight in modern birds are concentrated on the breast
(analogous to your pectoral muscles) which is why birds tend to have
really deep and round chests. These large muscles require an equally
large attachment, and in this case it’s the sternum, the solid
bone that connects the ribs together. If you have ever carved a
chicken or a turkey, then you will know that as you cut into the
breast you have to cut around a large flat bone that rises up from the
ribs; this is the sternum.
Evidence
for strongly developed flight muscles is mixed between specimens of
Archaeopteryx, but one thing for certain is that
there is no enlarged
bony sternum like that seen in modern birds. Some specimens currently
assigned to Archaeopteryx though show the possible
presence of a
cartilaginous sternum which would have functioned in the same way as a
bony one. Cartilage is a tissue that can be rigid in life, but
rarely survives long enough to fossilise except in cases where the
cartilage has become ossified (turned to bone) before death. Also
going back to carving chickens, while the main sternum is bone,
there is always a cartilaginous extension to this bone. If
Archaeopteryx possessed and enlarged cartilaginous
sternum, then they
may have been able to maintain a modified flight stroke that although
not as efficient as a modern bird, may have been able to allow for
sustained powered flapping flight. Otherwise all Archaeopteryx
would
be able to do was manage a few flaps of the wings before becoming tied
and having to stop, though this in itself might have been enough to
allow an individual to get airborne so that it could then glide the
rest of the way. It is also conceivable that from such a point as
this, later decedents that modified skeletons and musculatures could
eventually extend the length of time that they could keep flapping
until they evolved into birds more similar in form to what we know
today.
An
additional bit of support for flight in Archaeopteryx
actually comes from
analysis of the brain, specifically its size. Two studies by Witmer
and Alonso et al., and both in 2004 showed that relative to the
size of the body, the brain of Archaeopteryx was
actually quite
large. Reconstructions of the brain also indicated that the area
dedicated to vision accounted for one third of the total brain size,
while the areas of muscle co-ordination and hearing are also well
developed. The inner ear of Archaeopteryx is also
more like that of
modern birds that non-avian dinosaurs. What this all means is that
the brain of Archaeopteryx was already hardwired
with all of the
necessary features for controlling wings and moving in a three
dimensional environment like you would if you were flying.
When
you piece together the brain, the skeletal structure, the
musculature and feathers of Archaeopteryx, you
are left with a
creature that could definitely glide, and was perhaps capable of
limited powered flapping flight. In this respect Archaeopteryx
might
not have been a graceful flyer, but it possessed everything necessary
for further developments that appeared in later descendants. It
should always be remembered though that Archaeopteryx
is a transitional
form, if it were better developed for flight it probably would not be
so valuable for our understanding in bird evolution.
What did Archaeopteryx
look
like, and where did it live?
As
already mentioned, Archaeopteryx had a number of
dinosaur-like
features such as the arrangement of the shoulders, and the long
tail. The tail in modern birds is used to help with steering and
slowing down when descending, but the tail of Archaeopteryx
might not
have been that useful. The tail could have feasibly been used to
steer with minute adjustments, though its primary function may have
been that of a counterbalance while an Archaeopteryx
was on the
ground, just like with the theropod dinosaur ancestors.
The
flight feathers of the wings have already been mentioned, but smaller
downy feathers also covered the main body. So far no specimen has
shown the presence of feathers on either the head or the neck, but
there is actually uncertainty as to whether this was actually the case
in life. Up to 2013 all known specimens of Archaeopteryx
were
found in marine sediments, which means that they were exposed to
water before being buried in sediment. Outlined in a 2012 study
by Reisdorf and Wuttke, many dinosaurs are preserved in a
characteristic ‘death pose’ where the muscles and tendons of the
neck stiffen after death causing the head and neck to arc back, and
Archaeopteryx would have still been dinosaur-like
enough for this to
happen. This meant that as a body floated on the surface, the neck
and head would be the most underwater, causing the skin to soften and
the feathers to come loose giving the appearance of being bald.
Another study in 2002 by Elzanowski also suggested that the
feathers of the head and neck may have been worked loose as the body
rubbed against the sea bed before being buried.
The
colour of Archaeopteryx is not known for certain,
but a 2011
study by Carney et al. has given us a glimpse at what Archaeopteryx
might have been like. In this study a covert feather (a feather
that covers and protects others) from the wing was placed under an
electron microscope where melanosomes were discovered. A
melanosome
is basically a pigment, and when the melanosomes in the specimen were
compared next to those of 87 known bird species, the highest
likelihood was that in life the feather would have been black. This
means that the covert feathers on the wings of Archaeopteryx
were
possibly all black, but it is still unknown what colour the other
feathers were. They may of course have been black as well, but we
still do not know for certain.
Some
feathered dinosaurs and potential bird ancestors are better
understood. In the genus Anchiornis,
most of the feathers were
black, but white stripes ran down the wings, while red speckling
was on the face and an orange Mohawk style crown of feathers
was on top of the head. The dinosaur genus Sinosauropteryx
is
known to have had bands of deep brown and white running all the way
down its tail. We still do not know what the remainder of
Archaeopteryx looked like, but looking at other
genera tells us that
we should expect anything.
Archaeopteryx
is now known to have had three clawed fingers on each wing, and as
mentioned above, these may have been used to climb trees in a similar
way to juvenile Hoatzin. In Archaeopteryx though
there is no
indication that they were lost in adulthood, and were instead an
indication of dinosaurian heritage. The study of the Thermopolis
specimen (WDC CSG 100) in 2005 also revealed that
Archaeopteryx lacked a reversed toe, which means
that Archaeopteryx
could not grip hold of perches like modern birds. A hyperextensible
second toe was also found to be present, though this feature was
actually first conceived by Gregory S. Paul in 1988. The
hyperextensible second toe with claw and the three claws of the wings
may indicate that Archaeopteryx were
‘trunk-climbers’, climbing
to the trees instead of flying to them, but possibly jumping off and
gliding from tree to tree. Again this hints at the evolution in
flight for birds in that adaptations for gliding from tree to tree
leads to further adaptations for extended gliding ranges until full
powered flapping flight developed.
This
leads us to the ecosystems that Archaeopteryx lived
in. By 2013
all known specimens of Archaeopteryx are from
various locations in
Germany with the specimens from the Solnhofen limestone deposits being
the most famous. Central Europe during the Jurassic was actually a
series of islands surrounded by shallow seas and lagoons.
Rhamphorhyncoid pterosaurs were very common, but more advanced
pterodactyloid pterosaurs were also present. Many of the islands
seemed to have been quite arid, with some growths of cycads and
conifers. There were not a lot of opportunities for Archaeopteryx
to
climb up high, but then they might not have had to, the island
plants might have been large enough to provide cover and possibly be
used as gliding platforms. We still have much to learn about these
ancient island ecosystems, and the picture of their reconstructions
might change dramatically with future discoveries.
The
structure of the foot being more suited to ground locomotion than
grasping coupled with the still relatively long legs have been taken by
some as signs that Archaeopteryx may have been
cursorial, which means
that they spent their time on the ground instead of in the air. It
may be that Archaeopteryx climbed up into shrubs
and small trees for
night time protection from potential predators. Analysis of the
scleral rings of Archaeopteryx shows that they
match diurnal birds,
which means that Archaeopteryx would have only
been active during the
day time only.
Archaeopteryx
seems to have been fairly slow growing creatures that took two years
and eight months to grow to adult size. This is based upon a 2009
study by Erickson et al. which studied the growth stages of the
bones which also found that other primitive birds such as Jeholornis
and Sapeornis
also grew slowly, though others such as Confuciusornis
grew comparatively quickly. It may be that the slow growth of
Archaeopteryx may be a primitive trait that was sped
up in later bird
forms as their metabolisms altered.
So was Archaeopteryx
the first
and ancestor of birds?
Short
answer; probably not. The fossil record is always only ever going
to be a portion of what once lived, and will in that respect always
be incomplete. There are already other genera’s of animals similar to
Archaeopteryx that fall on either side of the
dinosaur/bird definition
line that may be older than Archaeopteryx. As
always in
palaeontology, it only takes one new fossil discovery to trigger a
change in an accepted theory, and with fossil genera being
discovered and named at a faster rate from so many more locations
around the world than ever before, this is more and more likely to
happen.
We
also cannot prove that Archaeopteryx was ‘the’
definitive ancestor
of modern birds, it is more likely just one of many forms that
existed during the late Jurassic, and if the development of birds can
be narrowed down to just one genus, then any one of those,
including Archaeopteryx could be the origin of the
genetic line to
modern birds. But you also need to be certain about how you define
ancestor. Do you use the first bird, the first theropod dinosaur,
reptile, amphibian, vertebrate fish or single celled organism?
Any one of them and an almost uncountable number more could be defined
as the ancestors of birds, it’s just down to where you choose to
begin.
This
is why palaeontologists tend to prefer to take the safer option of
regarding Archaeopteryx as a representative of the
form of an ancestral
candidate, rather than the actual ancestor, it allows for the genus
to be studied as it should, without requiring too much of a re-write
for later when new discoveries are made.
Further reading
- On the Archaeopteryx of Von Meyer, with a
description of the fossil
remains of a long-tailed species from the lithographic stone of
Solnhofen - Richard Owen - 1863.
- On the animals which are most nearly intermediate between birds and
reptiles - Thomas Henry Huxley - 1868.
- Remarks upon Archaeopteryx lithographica - Thomas
Henry Huxley - 1868.
- Further evidence of the affinity between the dinosaurian reptiles and
birds - Thomas Henry Huxley - 1870.
- Archaeopteryx and the origin of birds - J. H. Ostrom - 1976.
- Feathers of Archaeopteryx: Asymmetric vanes
indicate aerodynamic
function - A. Feduccia & H. B. Tordoff - 1979.
- Flight capability and the pectoral girdle of Archaeopteryx - S. L.
Olson & A. Feduccia - 1979.
- What size was Archaeopteryx? - D. W. Yalden - 1984.
- Archaeopteryx Relationship With Modern Birds -
Thomas Holtz Jr -
1995.
- A new genus and species for the largest specimen of Archaeopteryx
- Andrzej Elzanowski - 2001.
- Body plumage in Archaeopteryx: a review, and new
evidence from the
Berlin specimen - P. Christensen & N. Bonde - 2004.
- Structure and function of hindlimb feathers in Archaeopteryx
lithographica - N. Longrich - 2006.
- Was Dinosaurian Physiology Inherited by Birds? Reconciling Slow
Growth in Archaeopteryx - Gregory M. Erickson,
Oliver W. M. Rauhut,
Zhonge Zhou, Alan H. Turner, Brian D. Inouye, Dongyu Hu, Mark A. Norell
- 2009.
- Narrow Primary Feather Rachises in Confuciusornis
and Archaeopteryx
Suggest Poor Flight Ability. - Science. 328 (5980): 887–889. - Robert
L. Nudds & Gareth J. Dyke - 2010.
- Archaeopteryx was robed in black - Brian Switek -
2011.
- New evidence on the colour and nature of the isolated Archaeopteryx
feather - R. Carney, Jakob Vinther, Matthew D. Shawkey, Liliana d'Alba
& J�rg Ackermann - 2012.
- Re-evaluating Moodie's Opisthotonic-Posture Hypothesis in fossil
vertebrates. Part I: Reptiles - The taphonomy of the bipedal dinosaurs
Compsognathus longipes and Juravenator
starki from the Solnhofen
Archipelago (Jurassic, Germany) - A. G. Reisdorf & M. Wuttke -
2012.
- Synchrotron-based chemical imaging reveals plumage patterns in a 150
million year old early bird - Phillip L. Manning, Nicholas P. Edwards,
Roy A. Wogelius, Uwe Bergmann, Holly E. Barden, Peter L. Larson,
Daniela Schwarz-Wings, Victoria M. Egerton et al. - 2013.
- Archaeopteryx in 4D. - Journal of Vertebrate
Paleontology. 103. -
Ryan Carney, Julia Molnar, Earl Updike, William Brown, Jessie Jackson,
Matthew Shawkey, Johan Lindgren, Peter Sj�vall, Peter Falkingham
& Jacques Gauthier - 2014.
- Wing bone geometry reveals active flight in Archaeopteryx.
- Nature
Communications. 9 (1): 923. - Dennis F. A. E. Voeten, Jorge Cubo,
Emmanuel de Margerie, Martin R�per, Vincent Beyrand, Stanislav Bureš,
Paul Tafforeau & Sophie Sanchez - 2018.
----------------------------------------------------------------------------
Random favourites
 |
 |
 |
|