











































The Mosasaurs

The mosasaurs are one of the most spectacular success stories of the Mesozoic. In a period of time spanning less than 35 million years, they went from very small semi aquatic lizards to the apex predators of the world’s oceans. This success comes from the mosasaurs ability to rapidly adapt to a variety of aquatic habitats while at the same time attacking and hunting quite literally anything that was in the water with them. In all seriousness the only thing a big mosasaur needed to be concerned about was an even bigger mosasaur, which makes mosasaurs vicious, fascinating, terrifying and altogether awesome.
Early discoveries
Our
study into mosasaurs goes all the way back to 1764 when the first
ever mosasaur skull was discovered in the Netherlands. This skull was
fragmentary and initially thought to belong to a then unknown species
of fish. In the early 1770‘s a second partial skull was found.
After this a fellow by the name of Johann Leonard Hoffmann, a
retired army physician, discovered further fossil remains, and made
an effort to have them looked at by the wider scientific community.
Hoffman disagreed with the idea that these remains were of a fish and
instead thought they belonged to a gigantic crocodile. In 1786 a
Dutch professor named Petrus Camper came to the conclusion that the
fossils belonged to a toothed whale.
The
fossils were stored in the Fortress of Maastricht where they were kept
until the fortress fell to French military forces and the region of
Maastricht was annexed by the Napoleonic Empire. The fossils were
moved to France where Barth�lemy Faujas de Saint-Fond studied them and
came to agree with the idea that they belonged to a large crocodile.
However the original notes written by Petrus Camper had not gone to
France, and when Petrus’s son Adriaan Gilles Camper reviewed his
father’s notes he came to what would be remembered as a surprisingly
close discovery. Adriaan’s conclusion was that the fossils remains
were of a gigantic monitor lizard, and this is something that we now
know to have been as close to the right answer without actually
being right as you could get. Support for this idea came in 1808
when Georges Cuvier, a leading figure in the field of comparative
anatomy also agreed with the monitor lizard hypothesis.
It
was not until 1822 that these fossils were given the name
Mosasaurus
by William Conybeare, and the name which means ‘Meuse
River Lizard’ which was near where the fossils were located.
However at this time Mosasaurus was still
perceived to being a huge
monitor lizard. In 1829 Gideon Mantell (famous for naming
Iguanodon, the first plant eating dinosaur to be described to
science) made the name official when he designated the type species
as Mosasaurus hoffmanii in memory of Johann Leonard
Hoffmann (it
should also be noted that at this time many extinct animals that were
being named were often given genus names lacking a species name,
something that had to be sorted out by later naturalists and
palaeontologists).
It
was not 1854 that Mosasaurus was put forward as
an aquatic creature
that lived in the ocean. This came about from an observation by
Hermann Schlegel who noted that the limbs of Mosasaurus
were not suited
for land movement and would have been far more use as flippers. Later
discoveries would prove Schlegel right and by the closing decades of
the nineteenth century mosasaurs were not just being found in marine
fossil deposits of not just Europe but North America as well. By the
early twenty-first century mosasaur fossils have been recorded in Late
Cretaceous marine deposits on every major continent showing that the
mosasaurs were so successful that they spread out across the globe
wherever the habitat was suitable for them.
The Rise of the Mosasaurs
Most
of the known primitive mosasauroids are currently aged as living in the
Cenomanian and Turonian eras. The Cenomanian and the Turonian are the
first two time periods that make up the Late Cretaceous, and about
100.5 to 89.9 million years ago. What is interesting here is
that about the time of the boundary between the Cenomanian and Turonian
there was actually an anoxic event. An Anoxic event is where the
levels of dissolved oxygen in the water plummet, and the animals that
breathe through gills asphyxiate and die, making this an extinction
level event.
At
this time the main kinds of large predatory marine reptiles in the
water were ichthyosaurs,
plesiosaurs
and pliosaurs.
As reptiles
these were all air breathing creatures, so some may assume that a
drop in water oxygen levels would not affect them, and directly it
wouldn’t. Indirectly however this drop would have killed off colossal
amounts of fish and cephalopods (squids, ammonites, etc.),
which would have been the key prey species of the plesiosaurs,
ichthyosaurs and some pliosaurs. With far less prey to eat these
would die out through starvation, and indeed for the ichthyosaurs
things seem to have been particularly bad. Almost all of the
ichthyosaur genera that we know of that made it to see the Cenomaninan
of the Cretaceous vanished at about the time of this extinction event,
and the rare few that survived died out a little bit later in the
Cretaceous. The large Pliosaurs with genera such as Brachauchenius
that survived the extinction event itself disappear from the fossil
record in the Turonian, possibly as a result of the other marine
reptiles dying out making less prey available to them. Out of these
three groups only the plesiosaurs are known to have made it to the end
of the Cretaceous, and then seemingly in reduced numbers.
With
the oceans relatively empty of the classical large marine reptiles,
the stage was set for a new group of predators to arise. The early
and most primitive of mosasauroid forms such as Dallasaurus
and
Komensaurus
amongst others were well adapted to swimming, but they
were not yet suited to life in the open ocean. Their tails were
long and flattened for swimming, but the legs still capable of
walking on land. Fossils of these primitive mosasaurs were found in
what were shallow waters and coastal locations and this is most
probably the reason why these reptiles not only survived the
Cenomanian/Turonian extinction, but thrived during it.
Shallow
waters around the coast would be less susceptible to a drop in oxygen
levels of the open ocean as a greater abundance of aquatic coastal
plants and water turbulence from tides would increase oxygen levels
in these parts meaning that these areas would have been richer in
aquatic life than say the open ocean where many of the large marine
reptiles were used to doing their hunting. This in turn meant that
the coastal areas would have had still had a great abundance of prey
from fish, other kinds of cephalopods such as octopuses, as well as
various crustaceans like shrimps would have all been viable prey
options for the primitive mosasaurs. With the ability to still walk
about on land, primitive mosasaurs may have also been beach combers,
scavenging the dead bodies of other aquatic organisms such as fish and
maybe even fully aquatic marine reptiles like plesiosaurs and
ichthyosaurs.
Most
of the primitive mosasaurs were actually very small, most of the
currently known genera did not deviate much more than about a meter in
length. Again, this would have been to their benefit, because
smaller bodies don’t require so much food to keep going, so it is
easier to survive an extinction event. When fish levels began to rise
later in the Cretaceous, the mosasaurs seem to have started venturing
out further and further into the sea, until eventually they stopped
returning to the land all together. In the absence of competition
from other kinds of large marine reptiles, the mosasaurs began not
only growing bigger, but better adapted to fast stable swimming,
and high agility in the water. By the Santonian period, only
about 4 million years after the Cenomanian/Turonian extinction,
the mosasaurs had gone from being small one meter long semi-aquatic
lizards, to very big marine apex predators, that for the remainder
of the Cretaceous would continue to grow bigger and ever more
specialised.
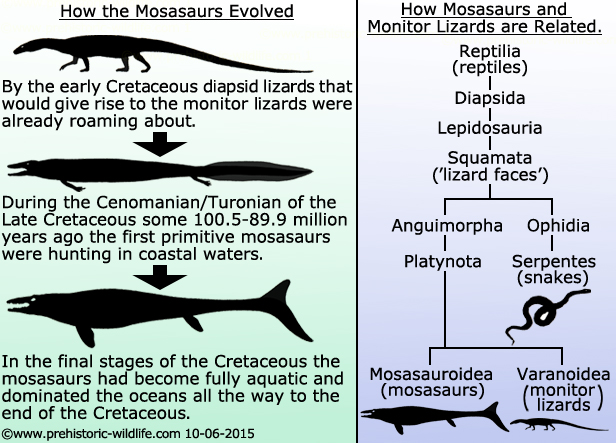
How are mosasaurs related to
snakes and monitor lizards?
For
a long time mosasaurs have been speculated to have been descended from
either monitor lizards or snakes, and it should be mentioned here
that both monitor lizards and snakes are also thought to have possibly
had a common ancestry. Mosasaurs had jaws that could open very wide
and were more suited to swallowing large chunks of prey, if not
actually swallowing their prey whole. Mosasaurs also had a second set
of teeth growing from their palate, and these teeth would help to
grip prey and larger chunks of flesh as they swallowed.
The
snake to mosasaur hypothesis is no longer that widely accepted. For a
start research and new snake fossil discoveries from the end of the
twentieth and early twenty-first centuries is already pointing to the
conclusion that while snakes and mosasaurs share some similarities,
these may actually be the result on convergent evolution, or shared
traits from a common but distant ancestor. Snakes are also known
to have only one functioning lung, the other being vestigial
(present but reduced to the point it does not work). One specimen
of the genus Platecarpus
has shown us that mosasaurs had two working
lungs, not one. Finally primitive mosasaurs forms such as
Dallasaurus
show us that the direct mosasaur ancestors had legs when
the adaptations were taking place.
Now
it seems that the original interpretation of Mosasaurus
being a monitor
lizard was actually only 40 million years off the mark. The
mosasaurs are now often regarded as a sister group to the Varanidae
which includes prehistoric monitor lizards such as Saniwa,
Palaeosaniwa
and the colossal Megalania/Varanus
priscus, as well as
the Varanus genus which includes all modern day
species of monitor
lizard.
The Various kinds of mosasaur
There
are two main family groups of mosasaur that you must be familiar with.
The first is the Mosasuroidea, which includes all mosasaurs,
including the primitive ones like Dallasaurus
and Komensaurus.
The
second group is the Mosasauridae, which includes all the advanced
form mosasaurs that are defined by Mosasaurus,
but excludes the
primitive forms.
The
Mosasauridae can be further divided into what are currently six
sub-families. First is the Mosasaurinae, which includes genera that
are physically close to Mosasaurus, which means
that regardless of
differences in size, are noted for having proportionately robust
builds meaning that members of the Mosasaurinae adapted to have an
emphasis on strength and power. Members of the Mosasaurinae could be
medium sized at around 6 to 7 meters long, to much larger at
12 to 15 meters long, with a few individuals exceeding even
this.
Second
is the Tylosaurinae, which includes the genera that are closer to
Tylosaurus.
So far tylosaurines and especially Tylosaurus
itself make
up in terms of body length some of the largest mosasaurs. However the
tylosaurines are notably more gracile (lightly built) than the
mosasaurines. This means that while tylosaurine mosasaurs were still
powerful, there emphasis was more on speed and agility than raw
strength. This means that a greater range of aquatic creatures was
within their predatory scope, and indeed stomach contents of
Tylosaurus do indicate that anything from fish to
other marine reptiles
to even sea birds were eaten by tylosaurine mosasaurs.
Third
is the Plioplatecarpinae, and this includes genera such as
Plioplatecarpus,
Platecarpus
and Angolasaurus
amongst many others.
These genera are notably smaller the previous two groups, with sizes
commonly ranging between 3 and 6 meters long.
Plioplatecarpines show some of the greatest adaptations for fast
swimming, with some genera developing adaptations that were analogous
to the earlier ichthyosaurs. It was the plioplatecarpine genus
Platecarpus that was the first to reveal the
presence of a fluked
tail, something that now seems to have been a common feature for
other mosasaurs. Plioplatecarpines also seem to have focused upon
swimming forward using only their tails instead of their whole bodies,
now an outdated idea.
Fourth,
fifth and sixth are the Halisaurinae, Yaguarasaurinae and
Tethysaurinae, and again these generally range from about 3 to
6 meters long. Like the Plioplatecarpine mosasaurs these seem to
have been predators of fish and possibly smaller marine reptiles. A
seventh group is called the Aigialosauridae after Aigialosaurus
but
this is a side group to the Mosasauridae which is a home for many of
the primitive mosasaurs.
Mosasaur biology
When
mosasaurs were first reconstructed as aquatic animals mosasaurs were
usually constructed as lizards with eel-like proportions. This idea
existed all the way into the twentieth century but eventually a
specimen of the mosasaur Platecarpus
was discovered and this included
partially preserved remains of soft tissue of the tail. This was the
first ever indicator that mosasaurs had bi-lobed tails with the main
tail dipping down and a soft tissue fluke rising up from the top. For
many though Platecarpus was the exception not
the rule, and so
reconstructions of larger mosasaurs usually went ahead with a more
classic tail.
Then
in 2008 a new specimen of the genus Prognathodon
was discovered,
and after five years of cleaning and study, it was revealed that a
second mosasaur was confirmed as having a bi-lobed tail with a fluke of
soft tissue rising up from the tail. Prognathodon
had a very
different ecological niche than what Platecarpus
had, and grew
significantly bigger, and this is the discovery that forced others to
sit up and think about how accurate the old reconstructions of
mosasaurs were. As of 2015, there is no hard evidence to support
the old hypothesis that mosasaurs had straight tails, but mounting
evidence that most if not all advanced mosasaurid mosasaurs did. For
this reason newer reconstructions of mosasaurs are always done with the
presence of a bi-lobed tail.
Mosasaurs
were air breathers which meant that they would periodically have to
swim to the surface and poke their head out of the water to fill their
lungs with fresh air. This was a very necessary but also dangerous
thing for mosasaurs to do, especially smaller genera which were
putting themselves at risk from attack from other larger mosasaurs as
well as some of the larger predatory sharks of the time such as
Cretoxyrhina,
Squalicorax
and Cardabiodon.
One
specimen of Platecarpus has been found with the
outlines and impression
of internal organs preserved on the slab of rock that the bones are
preserved upon. Here the trachea and bronchi are present, and show
that Platecarpus and probably other mosasaurs had
two working lungs.
Further to this when the bronchi split the trachea to take air into
each lung, the bronchi run parallel to each other instead of
branching out to the sides like in terrestrial lizards. This may show
an adaptation to the lungs working against the effects of water
pressure pushing in from the sides, as the same adaptation can be
seen in cetaceans (whales and dolphins) today.
It
is not improbable that mosasaurs may have a directional sense of
smell, something suggested by the confirmed placement of the nostrils
on Platecarpus which face out to the side. These
may have allowed for
water to be sampled, and if one nostril picked up a smell before the
other, a mosasaur might have been smart enough to figure out the
direction the smell, say something like blood from an injured animal
or a pheromone trace from a female ready to breed, and swim towards
it. Even animals that are not considered to be very intelligent can
do this, but monitor lizards which mosasaurs are close relatives of
are actually considered to be some of the most intelligent lizards.
There is not yet any way of knowing how smart a mosasaur was, but
the innate intelligence of monitor lizards may have been transferred to
mosasaurs and at least allowed them to develop much keener senses such
as sight and smell, as well as learned patterns of behaviour such as
knowing when certain kinds of animal would be available to hunt.
There is also fresh speculation that mosasaurs may have also had
forked tongues, though confirmed detail of this lacking at the time
of writing.
What
scales were on the body depends somewhat on the genus and body part in
question. Returning again to Platecarpus, there
seems to have been
three main kinds. On the actual body and underside of the tail the
scales are rhomboid and arranged into diagonal rows. Each row also
overlaps the row behind it, perhaps to increase hydrodynamic
efficiency. The upper side of the tail were similar to the rest of
the body but larger in size, perhaps as a way to increase the tails
rigidity for more efficient movement. On the snout of Platecarpus
however scales were hexagonal and not shaped in any regular
pattern, perhaps as a result of contact damage with prey animals.
In specimens of other mosasaurs scales on the upper body were
keeled, perhaps again to increase rigidity, or perhaps even to
prevent light scattering so that they could more easily lurk in the
murky depths and wait for their target prey to return to the surface
for fresh air where they would be most vulnerable.
For
a long time the colour of mosasaurs was unknown, and this was true
until 2014 when a paper (Lindgren et al.) described the process
of examining melanin pigments in a sample of fossilised mosasaur skin.
The result for the specimen in question was that mosasaurs like a
great many other marine creatures were counter shaded, with very dark
almost black colouration on the sides and back, while very lightly
coloured on the underside. It is not yet known if all mosasaurs
followed suit, indeed species specialising in hunting in shallows may
have been quite different. But most pelagic (open water) animals
do have this patterning, and if one mosasaur did, then it’s
reasonable that at least many others also did.
Mosasaurs
are confirmed as being viviparous, which means that mosasaurs did not
lay eggs but gave birth to live young. This is not that farfetched as
while reptiles are generally associated with laying eggs, some
lizards and snakes have been observed and scientifically documented to
give birth to live young. In mosasaurs direct evidence from this
comes from the genus Clidastes
which had a much smaller juvenile
Clidastes inside of it. Remains of small juveniles
have been found as
stomach contents before, but these are always partially corroded by
acid, with distinctive pitting in the bones being evident of contact
with the stomach acid. The small Clidastes in the
larger one is in
pristine condition meaning no exposure to stomach acid. This raises
the likelihood that the adult Clidastes was
pregnant at the time of
death, and confirms an idea that mosasaurs did not lay eggs on land
because of the fantastically great difficulties they would have had
trying to crawl out of the water.
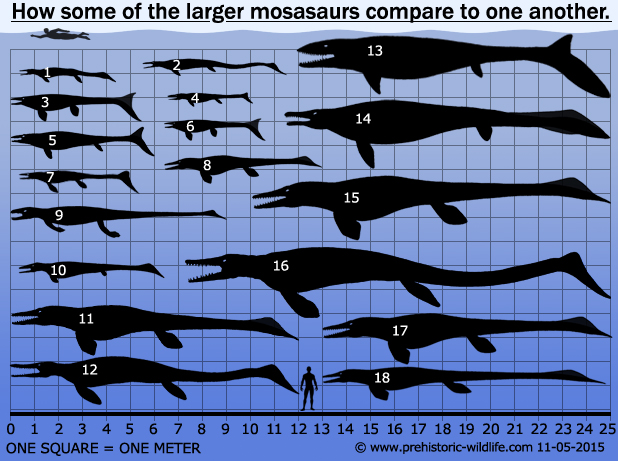 |
| 1 - Halisaurus, 2 - Pannoniasaurus, 3 - Plioplatecarpus, 4 - Carinodens, 5 - Globidens, 6 - Platecarpus, 7 - Plesioplatecarpus, 8 - Plesiotylosaurus, 9 - Yaguarasaurus, 10 - Clidastes, 11 - Hainosaurus, 12 - Liodon, 13 - Prognathodon, 14 - Plotosaurus, 15 - Tylosaurus, 16 - Mosasaurus, 17 - Taniwhasaurus, 18 - Moanasaurus. |
How did mosasaurs swim?
A
long standing idea since the early days of
mosasaur research is that mosasaurs swam but undulating their entire
bodies from side to side like a snake does. There are three reasons
why this idea is no longer thought sound. First is that mosasaurs are
probably not related from snakes, so there is no reason to assume
that they would swim like them. Second is that such a method of
swimming is relatively slow, and would be a question rather than an
answer regarding how mosasaurs became so successful. Third, modern
analysis of mosasaur bodies and scale patterning shows that when
swimming at least the body would have been held fairly rigid to reduce
resistance when moving through the water.
This
means that the only effective method of propulsion of swimming would be
to move the tail, and the tail only from side to side. The presence
of a tail fluke which is now widely accepted as a fact for most if not
all mosasaurs would have greatly increased the amount of push the tail
had against the water making faster speeds more easily possible, as
well as bursts of quick speed when attacking prey. Scales of
mosasaurs when preserved are often fairly small but also shaped to
decrease drag, making it easier for a mosasaur to move through the
water with the least amount of effort.
One
myth about mosasaurs swimming which is still put forward even in modern
times is that mosasaurs would use their flippers for actual
propulsion. This is just plain wrong, as moving the flippers in
conjunction with the tail would only increase drag, reduce speed and
increase the amount of energy required for swimming. The true
function of the flippers was far simpler: steering. The problem
with tails is that they push the body forward, not pull. This means
that the body is always ‘nose heavy’ and will pitch down from
gravity plunging the body towards the bottom of the ocean. The front
flipper at least of a mosasaur would have been held at to the sides
during regular everyday cruising and angled to act as hydroplanes to
counteract this pitching effect.
This
is actually a sound scientific principal, all fish use their pectoral
fins at the front to counter this pitching effect, even submarines
built by mankind have bow planes to counter the same effect. The
flippers on mosasaurs could have also been used for steering allowing
mosasaurs to make what were relatively tight turns in the water making
them allow the more precise and deadly predators. When making a final
attack however, mosasaurs may have folded their flippers back as
close to their body as possible to reduce drag and increase their
attack speed as much as possible, giving their prey as small as
possible a chance to escape as they could.
Where did mosasaurs live?
Mosasaurs
could live anywhere they had water to swim in. As air breathers it
did not matter if they were in either salt water or fresh water. The
overwhelming majority of mosasaur fossils are known from marine (salt
water) sediments, and even more often from areas that were once
shallow seas or coastlines. However, while the sea was undoubtedly
their favourite haunt, the holotype fossils of the genus
Pannoniasaurus and Goronyosaurus
were recovered from what are percieved to have once been fresh water
areas. It would be impossible for a dead mosasaur to float
upstream, rivers always flow into the sea, so this can only mean
that this individual made the decision to swim upstream. Since
mosasaurs gave birth to live young, it is perhaps then more likely
that Pannoniasaurus and Goronyosaurus
were on the hunt. There is
absolutely no
reason to assume that other mosasaurs did not swim into river systems,
and while there may have had a predator/prey interaction with some
dinosaurs.
As
far as geographic distribution goes, mosasaurs are known from every
major continent in the world, even Antarctica (which in the
Cretaceous was further North and warmer than what it is today).
The hotspots for mosasaur fossils however are currently Europe,
North Africa and the central portion of North America, with a
special focus on the central states of the USA. This is because in
the Late Cretaceous continental Europe was more like a chain of islands
surrounded by shallow seas, North Africa was surrounded by shallow
seas and had extensive rivers systems, and the central area of
North America was submerged by the western interior seaway. This area
were near perfect for preserving the remains of mosasaurs, and are
also known for producing large numbers of fossils for other marine
animals.
This
is not to suggest that mosasaurs were more common in these areas during
the Late Cretaceous, we can only study marine sediments from that
time, and if they have not been preserved there is not much we can do
further. Given their ancestry however mosasaurs were probably
ectothermic, meaning that they were more reliant upon the temperature
of the water to keep their bodies at the right temperature. For this
reason the mosasaurs may have favoured waters in the warmer latitudes,
which in the Late Cretaceous were more widespread given that global
temperatures seem to have been higher.
What did mosasaurs eat?
In
the simplest terms mosasaurs would eat anything that they could catch.
Tylosaurus
is the best example of this with fossils of animals found
partially preserved in what would have been the stomach including
remains of fish, sharks, marine reptiles, including other
mosasaurs and even sea birds such as Hesperornis.
It was their
voracious appetites that allowed mosasaurs to be so successful,
absolutely nothing in the ocean would be safe from an attack by a
mosasaur.
The
largest of the mosasaurines were certainly the apex predators that
would focus their hunting upon other mosasaurs and plesiosaurs, as
well as possibly larger sharks and fish. The smaller and faster
mosasaurs were undoubtable hunters of fish, either cruising over
shallow reefs and snatching out fish that darted out from amongst the
rocks, or perhaps harassing large fish shoals in the open water.
The
mosasaurines with their more robust skulls and heavier builds could
have still attacked marine reptiles like plesiosaurs and mosasaurs that
were on the surface, but they would have also been better suited to
attacking more heavily armoured prey like the large protostegid
turtles, typified by such genera as Archelon
and Protostega.
Two
mosasaurine genera in particular though, Prognathodon
and
Globidens
had specially adapted teeth that would easily break up the shells of
turtles and ammonites.
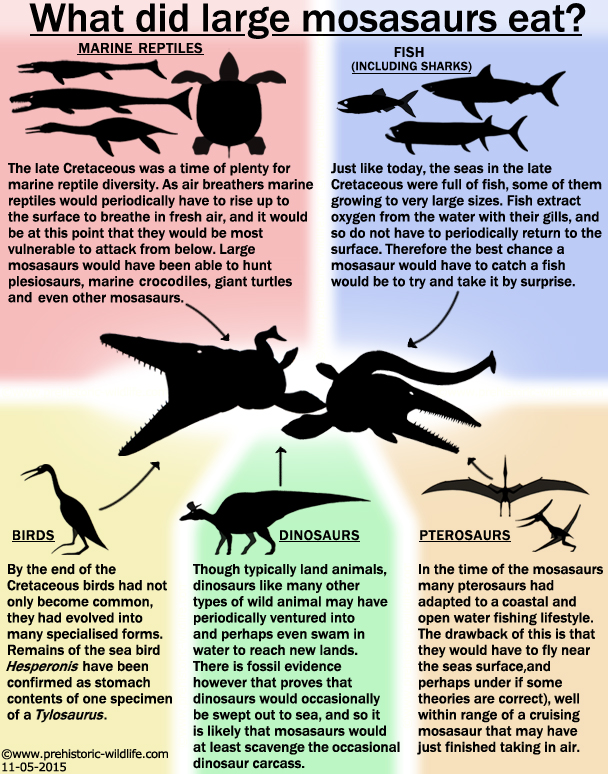
Extinction
Just
as an extinction event may have helped clear the seas for mosasaurs to
rise to power, an even greater event, the KT extinction marks their
disappearance from the fossil record. The KT Extinction marks not
only the end of the Maastrichtian and the Cretaceous, but the
Mesozoic as well, and the mosasaurs along with the plesiosaurs,
pterosaurs, and even the dinosaurs all disappear. This is event is
signified by the famous asteroid strike that took place in the north of
the Yucatan Peninsula, however new and ongoing research suggests that
there may have been more than one major asteroid strike responsible.
In addition to this volcanic activity may have been much greater than
what we know today, and these factors combined may have been what
wiped out the mosasaurs as well as many other types of Mesozoic animals.
Although
different mosasaurs had very different diets to one another, they
were all part of a global food web, and one upset, particularly at
the most basic level would have larger ramifications for all of the
other creatures concerned. If one or more major asteroid strikes
occurred or massive volcanic eruptions continued to happen, or even a
combinations of these all at once, then the skies would have become
darkened as ash and dust were ejected into the atmosphere. This would
reduce the rate of photosynthesis causing a large scale reduction in
the amount of plankton. This would reduce available food to marine
organisms like shrimps and small fish that ate the plankton, which
would reduce the numbers of bigger fish that ate them. This would
then in turn reduce the numbers of their predators such as the smaller
mosasaurs and plesiosaurs, which in turn would reduce the numbers of
their predators, the larger mosasaurs. As unfavourable conditions
to life continued, the declines would continue until the population
levels reached a point that a species could not some back from, then
it would only be a matter of time before that species went extinct.
In
addition to that, large amounts of dust and ash in the atmosphere
would block out sunlight and reduce global temperatures, even the
temperatures of the oceans. If ectothermic and dependent upon the
water temperature to maintain a good metabolism, then this would have
had a further negative impact on the mosasaurs, making them more
sluggish and less able to hunt, as well as maybe even reducing their
ability to reproduce. This is because many reptiles that we know
today will not even begin to try and reproduce until they have been
exposed to an ‘ideal’ temperature for a period of time.
After
the mosasaurs went extinct the world’s oceans were relatively empty
of impressively large apex predators. The sharks survived the
extinction, biologically speaking sharks were and continue to be far
better adapted to life in the oceans than any marine reptile. However
it would not be until the mammals started venturing into the water in a
similar way that the first primitive mosasaurs did that the world’s
oceans would begin to see gigantic predators again. In an
evolutionary pattern that would repeat itself, the first ancestors of
the whales were beach combers that would occasionally venture out into
the water to reach new areas and hunt for animals in the water. By
the Eocene period whales were living in the water permanently, and
had developed long streamlined bodies for swimming into the water. In
fact some of these primitive whales were so similar to the mosasaurs
that one genus named Basilosaurus
was actually described as a mosasaur
before it was realised to be a whale. It is also the rise of these
whales that would see some gigantic sharks such as C.
megalodon
appear to hunt them.

List of some mosasaur genera
? - Denotes a genus that has been named but the validity of which has now been questioned by some palaeontologists.
Further reading
- Notes on remains of fossil reptiles discovered by Prof Henry Rogers
of Pennsylvania, US, in Greensand Formations of New Jersey. - Quarterly
Journal of the Geological Society of London 5(1):380-383. - Richard
Owen - 1849.
- A Complete Mosasaur Skeleton, Osseous and Cartilaginous. - Memoirs of
the American Museum of Natural History 1 (4): 167–188. - Henry
Fairfield Osborn - 1899.
- A Mounted Skeleton of Platecarpus. - The Journal of Geology vol 18 -
S. W. Williston - 1910.
- An aberrant mosasaur from the Upper Cretaceous of north-western
Nigeria. - Rendiconti della classe di fisiche, mathematiche e naturali
52:398-402. - A. Azzaroli, C. De Giuli, G. Ficcarelli & D.
Torre - 1972.
- Tooth morphology and prey preference of Mesozoic marine reptiles. -
Journal of Vertebrate Paleontology 7 (2): 121–137. - J. A. Massare -
1987.
- The phylogeny of varanoid lizards and the affinities of snakes. -
Philosophical Transactions Royal Society London B Biological Sciences.
352 (1349): 53–91. - M. S. Y. Lee. - 1997.
- Live birth in Cretaceous marine lizards (mosasauroids). Royal society
proceeding B vol 268 - Michael W. Caldwell & Michael S. Y. Lee
- 2001.
- Molecular evidence for a terrestrial origin of snakes. -
Philosophical Transactions Royal Society London B Biol Sciences. 271:
S226–S229. - N. Vidal & S. B. Hedges - 2004.
- Durophagous Mosasauridae (Squamata) from the Upper Cretaceous
phosphates of Morocco, with description of a new species of Globidens -
Netherlands Journal of Geosciences - Geologie en Mijnbouw
84(3):167-175. - N. Bardet, X. Pereda Suberbiola, M. Iaroch�ne, M.
Amalik & B. Bouya - 2005.
- Dallasaurus turneri, a new primitive mosasauroid from the Middle
Turonian of Texas and comments on the phylogeny of the Mosasauridae
(Squamata). - Netherlands Journal of Geoscience (Geologie en Mijnbouw)
84 (3) pp. 177-194. - G. L. Bell Jr. & M. J. Polcyn - 2005.
- A new species of Halisaurus from the Late Cretaceous phosphates of
Morocco, and the phylogenetical relationships of the Halisaurinae
(Squamata: Mosasauridae). - Zoological Journal of the Linnean Society
143:447-472. - N. Bardet, X. Pereda Suberbiola, M. Iarochene, B. Bouya
& M. Amaghaz - 2005.
- A Cretaceous terrestrial snake with robust hindlimbs and a sacrum. -
Nature 440 (7087): 1037–1040. - S. Apestegu�a & H. Zaher - 2006.
- Latest Cretaceous mosasaurs and lamniform sharks from Labirinta cave,
Vratsa District (northwest Bulgaria): A preliminary note. - Geoloski
anali Balkanskoga poluostrva 67: 51–63. - John W.M. Jagt, Neda
Motchurova-Dekova, Plamen Ivanov, Henri Cappetta & Anne S.
Schulp - 2006.
- A new basal mosasauroid from the Cenomanian (U. Cretaceous) of
Slovenia with a review of mosasauroid phylogeny and evolution. -
Journal of Vertebrate Paleontology 27(4):863-880. - M. W. Caldwell
& A. Palci - 2007.
- A redescription of Aigialosaurus (Opetiosaurus) bucchichi (Kornhuber,
1901) (Squamata: Aigialosauridae) with comments on mosasauroid
systematics. - Journal of Vertebrate Paleontology 29 (2): 437-452. -
Alex R. Dutchak & Michael W. Caldwell - 2009.
- On the latest scale coverings of mosasaurs (Squamata: Mosasauridae)
from the Harrana Fauna in addition to the description of s new species
of Mosasaurus. - Fossils of the Harrana Fauna and the Adjacent Areas.
Amman: Eternal River Museum of Natural History. pp. 80–94. - H. F.
Kaddumi - 2009.
- Redescription of the holotype of Platecarpus tympaniticus Cope, 1869
(Mosasauridae: Plioplatecarpinae), and its implications for the alpha
taxonomy of the genus. - Journal of Vertebrate Paleontology
30(5):1410-1421- T. Konishi, M. W. Caldwell & G. L. Bell Jr. -
2010.
- Convergent evolution in aquatic tetrapods: insights from an
exceptional fossil mosasaur. PLoS One 5(8):e11998 - J. Lindgren, M. W.
Caldwell, T. Konishi & L. M. Chiappe - 2010.
- Convergent Evolution in Aquatic Tetrapods: Insights from an
Exceptional Fossil Mosasaur. - PLoS ONE 5 (8): e11998. - J. Lindgren,
M. W. Caldwell, T. Konishi & L. M. Chiappe - 2010.
- Microspectroscopic Evidence of Cretaceous Bone Proteins. - PLoS ONE 6
(4). - J. Lindgren, P. Uvdal, A. Engdahl, A. H. Lee, C. Alwmark,
Karl-Erik Bergquist, Einar Nilsson, Peter Ekstr�m, Magnus Rasmussen,
Desir�e A. Douglas, Michael J. Polcyn & Louis L. Jacobs - 2011.
- New exceptional specimens of Prognathodon overtoni (Squamata,
Mosasauridae) from the upper Campanian of Alberta, Canada, and the
systematics and ecology of the genus. - Journal of Vertebrate
Paleontology vol 31, issue 5 - T. Konishi, D. Brinkman, J. A. Massare
& M. W. Caldwell - 2011.
- A combined evidence phylogenetic analysis of Anguimorpha (Reptilia:
Squamata). - Cladistics 27 (3): 230–277. - Jack L. Conrad, Jennifer C.
Ast, Shaena Montanari & Mark A. Norell - 2011.
- A Large Mosasaurine (Squamata: Mosasauridae) from the Latest
Cretaceous of Osaka Prefecture (Sw Japan). - Paleontological Research
16 (2): 79–87. - Takuya Konishi, Masahiro Tanimoto, Satoshi Utsunomiya,
Masahiro Sato & Katsunori Watanabe - 2012.
- The first freshwater mosasauroid (Upper Cretaceous, Hungary) and a
new clade of basal mosasauroids. - PLoS ONE 7(12):e51781. - L. Mak�di,
M. W. Caldwell & A. Osi - 2012.
- Mass extinction of lizards and snakes at the Cretaceous–Paleogene
boundary. - Proceedings of the National Academy of Sciences of the
United States of America 109 (52): 21396–21401. - Nicholas R. Longrich,
Bhart-Anjan S. Bhullar and Jacques A. Gauthier - 2012.
- Soft tissue preservation in a fossil marine lizard with a bilobed
tail fin. - Nature Communications 4. - J. Lindgren, H. F. Kaddumi
& M. J. Polcyn - 2013.
- Reassessment of Turonian mosasaur material from the 'Middle Chalk'
(England, U.K.), and the status of Mosasaurus gracilis Owen, 1849. -
Journal of Vertebrate Paleontology 34 (5): 1072–1079. - Hallie P.
Street & Michael J. Caldwell - 2014.
- Skin pigmentation provides evidence of convergent melanism in extinct
marine reptiles. - Nature 506 (7489): 484–8. - J. Sj�vall, P. Sj�vall,
R. M. Carney, P. Uvdal, J. A. Gren, G. Dyke, B. P. Schultz, M. D.
Shawkey, K. R. Barnes & M. J. Polcyn - 2014.
- Pelagic neonatal fossils support viviparity and precocial life
history of Cretaceous mosasaurs. - Palaeontology vol58 issue 3
pp401-407. - Daniel J. Field, Aaron LeBlanc, Adrienne Gau &
Adam D. Behlke - 2015.
----------------------------------------------------------------------------
Random favourites
 |
 |
 |