












Saurolophus
Name:
Saurolophus
(lizard crest).
Phonetic: Sore-o-loe-fus.
Named By: Barnum Brown - 1912.
Classification: Chordata, Reptilia, Dinosauria,
Ornithischia, Ornithopoda, Hadrosauridae, Saurolophinae.
Species: S. osborni
(type), S. angustirostris. Other species
sometimes mentioned are S. "morrisi" and S.
kryschtofovici, but more on these are in the main text.
Diet: Herbivore.
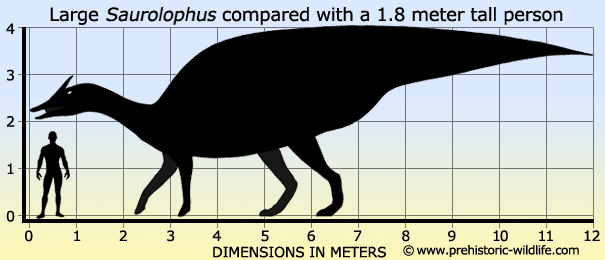
Size: Between 10 and 12 meters long,
depending upon species.
Known locations: Canada - Alberta -
Horseshoe Canyon Formation. Mongolia - Nemegt Formation.
Possibly also USA - California - Moreno Formation and China
- Heilongjiang Province.
Time period: Early Maastrichtian of the Cretaceous.
Fossil representation: Many skulls and skeletons,
some of which are almost complete. Skin impressions are also known.
Although
often labelled as a duck billed dinosaur, Saurolophus
is more
technically defined as a hadrosaurid
due to its placement within the
Hadrosauroidea. Even more specifically, Saurolophus
is defined as
the type genus of the Saurolophinae, the group of solid crested to
crestless hadrosaurids that were previously known as hadrosaurines
before the then type genus of Hadrosaurus
was found to not actually be
a member of this group.
Saurolophus
fossils were first discovered in Alberta, Canada along the Red Deer
River. Used to establish the type species of S. osborni
by Barnum
Brown, S. osborni may have shared its
environment with other
hadrosaurid genera such as Edmontosaurus
and Hypacrosaurus.
Compared
to the known remains of these other two genera however, Saurolophus
does not seem to have been as common. Here in Alberta at this time,
Saurolophus and other dinosaurs like it may have
been preyed upon by
large tyrannosaurs
such as Albertosaurus,
though smaller theropods
like Troodon,
Saurornitholestes
and Richardoestesia
may have also
been threats, particularly to smaller juvenile Saurolophus.
S.
osborni would have likely come into contact with other kinds
of
dinosaurs such as horned ceratopsians
like Pachyrhinosaurus
and
Anchiceratops,
as well as armoured ankylosaurs like nodosaurs like
Euoplocephalus
and Edmontonia
respectively.
The
species that has really increased out understanding of Saurolophus
however is S. angustirostris (named by
Rozhdestvensky, 1952),
based upon fossils from the Nemegt Formation of Mongolia. There are
subtle differences in the skulls of these species which in S.
angustirostris is twenty per cent longer than S.
osborni. Skin
impressions for both these species are also known and again there
appears to be a difference in the scalation. S.
angustirostris had
rows of square scales that ran along the back and tail, as well as
having varying scales forming vertical stripes down the tail. By
contrast S. osborni lacked the square scales
along its back,
instead being smooth, and the scales on the tail formed radial
patterns instead of vertical stripes. The most obvious difference
between these species however is that at twelve meters long S.
angustirostris seems to have been much larger than S.
osborni which
is so far only known from fossils that point to a length of just under
ten meters long.
Aside
from these differences, S. angustirostris seems
to have lived in a
similar way to S. osborni, though the
significantly higher number
of fossils attributed to this species suggest that S.
angustirostris
was one of the more common herbivores in its ecosystem. Like with S.
osborni, S. angustirostris likely
came into
contact with
ankylosaurs though this time genera like Tarchia, but higher levels
of other kinds of dinosaurs such as ornithomimosaurs
like Gallimimus
and the mysterious Deinocheirus,
oviraptorids like Avimimus,
therizinosaurs like Therizinosaurus
and probably most interestingly
sauropods like Nemegtosaurus
and Opisthocoelicaudia.
Again principal
predators of S. angustirostris would have
included tyrannosaurs,
though this time Tarbosaurus
and Alioramus.
Smaller predators such
as Zanabazar
and Adasaurus
could have also been a threat.
Another
species called S. kryschtofovici (named by
Riabinin, 1930) was
based upon the description of an incomplete ischium (part of the
hip) recovered from Heilongjiang Province of China. Due to the
fragmentary nature of the fossil it is based upon, S.
kryschtofovici is widely considered a nomen dubium because
it tis near
impossible to attribute any further fossil discoveries to this
species. Because the ischium in question was discovered in China,
there has been light speculation that it might actually be a specimen
of the better known S. angustirostris from nearby
Mongolia. If a
link could be established then under standard rules governing the
naming of animals, S. angustirostris would
actually become a junior
synonym to S. kryschtofovici since this species
was named first.
However this is unlikely to happen because of the aforementioned poor
state of preservation for the S. kryschtofovici
type specimen.
A
possible species, the description for which is still in press at the
time of writing is S. morrisi. These remains
were first discovered
in the Moreno Formation of California, USA between 1939 and
1940 but not assigned to Saurolophus (with
at least one skull to
Edmontosaurus) until 2010. Later study saw a
more complete
description of the remains as S. morrisi by
Prieto-M�rquez &
Wagner in 2012. One significant thing about this discovery is that
these are the first dinosaur remains discovered in the Moreno Formation
which is actually full of marine reptiles. Some of these include
mosasaurs like Plotosaurus
to plesiosaurs like Aphrosaurus
and
Morenosaursus,
so it will be interesting to see how dinosaurs like
Saurolophus existed in these ecosystems,
especially when you consider
that others such as Lambeosaurus
are also thought to have had a
particularly close association with marine ecosystems.
The
distinguishing feature of Saurolophus is the crest
which forms a single
blunt spike-like rearward projection that rises from the top of the
skull. Interestingly for a saurolophine hadrosaurid, this crest is
not completely solid, and is actually hollow where it joins the
skull. Despite this Saurolophus is still
classified with the other
solid/non-crested hadrosaurids because of similarities in the hips
between these genera, as well as a crest that while partly hollow,
is still more solid than the hollow crested lambeosaurine hadrosaurids.
Like
with other hadrosaurids, Saurolophus is thought
to have been
primarily quadrupedal in standard locomotion, but possibly bipedal
when foraging or possibly even running from predators. By being able
to switch between bipedal and quadrupedal postures, Saurolophus
was
capable of feeding upon plants of varying heights, from low down near
the ground to moderately high beyond the range of other low
browsers. The ‘duck-bill’ of Saurolophus
would have been
effective at snipping vegetation so that mouthfuls could easily be
passed back to the rear grinding teeth. A theory that is steadily
gaining more popularity is that hadrosaurs like Saurolophus
where able
to ‘chew’ because of special skull bones that came apart when the
lower jaw was pressed against them. This would cause the upper and
lower batteries of grinding teeth to slide against each other like two
pieces of sandpaper to process vegetation before swallowing.
The
eyes of Saurolophus were supported internally by
bony growths called
scleral rings that helped keep the shape of the eye. These rings are
also known in modern day reptiles and birds, and when the scleral
rings of Saurolophus are compared with species that
we can observe
today we notice that they most closely match those of cathemeral
reptiles and birds. This means that Saurolophus
was likely active for
short periods throughout the day, during both daylight and night-time
hours. This also matches theories of about the daily activity
patterns of other hadrosaurid genera where the scleral rings are also
known and found to be cathemeral also.
Further reading
- A crested dinosaur from the Edmonton Cretaceous, Barnum Brown
- 1912.
- The skeleton of Saurolophus, a crested
duck-billed dinosaur from
the Edmonton Cretaceous, Barnum Brown - 1913.
- [A new representative of the duck-billed dinosaurs from the Upper
Cretaceous deposits of Mongolia], Anatoly K. Rozhdestvensky -
1952.
- Cranial anatomy of Saurolophus angustirostris
with comments on the
Asian Hadrosauridae (Dinosauria), Teresa Maryanska &
Halszka Osmolska - 1981.
- Revision of the status of Saurolophus
(Hadrosauridae) from
California, USA, P. R. Bell & D. C. Evans -
2010.
- Saurolophus morrisi, a new species of
hadrosaurid dinosaur from
the Late Cretaceous of the Pacific coast of North America, Albert
Preito-Marquez & Jonathan R. Wagner - 2011.
- Cranial osteology and ontogeny of Saurolophus
angustirostris from
the Late Cretaceous of Mongolia with comments on Saurolophus
osborni
from Canada, Phil R. Bell - 2011.
- Nocturnality in Dinosaurs Inferred from Scleral Ring and Orbit
Morphology, L. Schmitz & R. Motani - 2011.
- Standardized Terminology and Potential Taxonomic Utility for
Hadrosaurid Skin Impressions: A Case Study for Saurolophus from Canada
and Mongolia. - PLoS ONE. 7 (2). - P. R. Bell - 2012.
- Perinatal specimens of Saurolophus angustirostris (Dinosauria:
Hadrosauridae), from the Upper Cretaceous of Mongolia. - PLOS ONE. 10
(10). - Leonard Dewaele, Khishigjav Tsogtbaatar, Rinchen Barsbold,
G�raldine Garcia, Koen Stein, Fran�ois Escuilli� & Pascal
Godefroit - 2015.
Random favourites
 |
 |
 |
 |